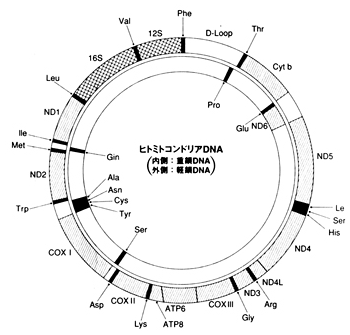
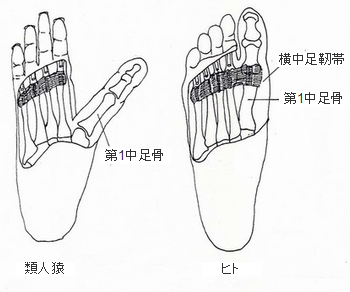
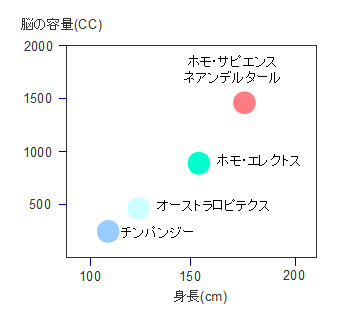
�z����T�s�G���X�̊g�U
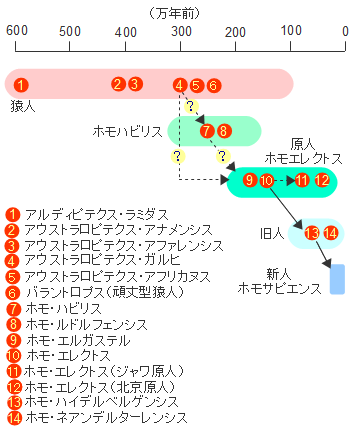
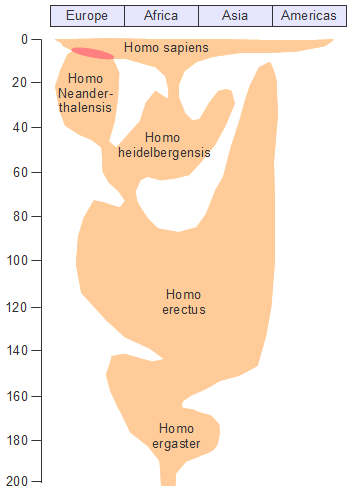
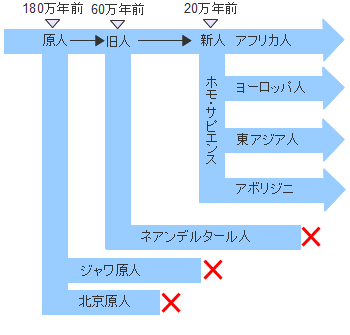
�A�t���J�P��N�����ł͌����l�ނ͂��ׂăz���E�T�s�G���X�ł���C6-10���N�O�ɃA�t���J�嗤���o�Đ��E�e�n�Ɋg�U���Ă����܂����B�܂��C�A�t���J�嗤����o���̂�1�x�������Ƃ�����肪����܂��B�~�g�R���h���ADNA�̕��͌��ʂ͌���ɂȂ���n���̓A�t���J���o����̏W�c�ł��邱�Ƃ��������Ă��܂��B�������C����قǒP���Șb�ł͂Ȃ���������܂���B
���Ƃ��C���������߂��W�c�̑O�ɂ��A�t���J���o���W�c������C�ނ�͊e�n�ɐ������Ă�����������܂���B�������C�ォ�����Ă��������W�c�Ɉ��ݍ��܂�Ă��܂��C���ʓI�Ƀ~�g�R���h���ADNA�ɂ͎c��Ȃ������Ƃ����\�����ے�ł��܂���B
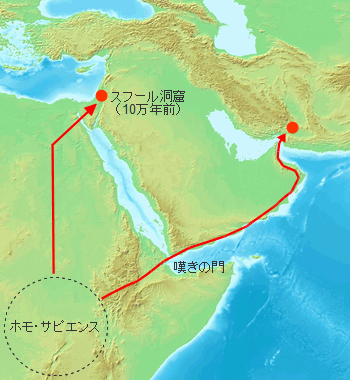
���ݔ�������Ă���ŌẪz���E�T�s�G���X�̉����̓G�`�I�s�A�Ŕ������ꂽ19.5���N�O�̂��̂ł��B16.5���N�O�ɂ͓�A�t���J��[�̊C�ݓ��A�ŕ�炵�����Ղ��������Ă��܂��B�ނ�͐��I�ȍאn�̐Ί���g�p���C�C�Y�����H�ׂĂ������Ƃ��������Ă��܂��B�C�Y���̗��p�͓��H�����Ƃ̋������Ȃ��s���̂悢�H���ł������ƍl�����܂��B
�܂��C�I�[�J�[�Ƃ���������Ċ痿������Ă������Ղ�����܂��B�痿�͊�G�Ɏg�p���ꂽ��C�̂ɓh�����肵�Ă����ƍl�����܂��B�z���E�T�s�G���X�͂��̏����i�K���當���I�ȖG�肪�������悤�ł��B
�A�t���J���C�݂̃z���E�T�s�G���X�̍��Ղ���͎����͓����ł͂���܂��C�痿�C�r�[�Y�Ȃǂ̑����i�C�ߗނȂǂ���������Ă��܂��B�܂�C�z���E�T�s�G���X�̓A�t���J���o��ȑO���������̌��^�������Ă����Ƃ������ƂɂȂ�܂��B
�����҂ɂ���ẮC�����Ԍ𗬂̂Ȃ������암�A�t���J�Ɠ��A�t���J�̌���l�W�c�̒��ɗގ���������������̂����邱�Ƃ���C�z���E�T�s�G���X�̑����i�K�������ɋ߂����̂����łɂ������Ƃ���l�����܂��B
���Ă̊w���ł̓��[���b�p�ɂ�����I�[���j���b�N�����i4��-1.5���N�O�j���z���E�T�s�G���X�ɂ�镶���̎n�܂�ł���Ƃ��Ă��܂����C�����̌��^�͂��łɃA�t���J�łł��Ă����Ƃ��������́C�ނ�̃v���C�h��������Ə�������������܂���B
�A�t���J�O�̃z���E�T�s�G���X�̍ŌẨ����̓C�X���G���̃X�t�[�����A�Ō�������10���N�O�̂��̂ł��B12���N�O�͐���N�Ԃ̊ԕX��������C�T�n���ɂ��ꎞ�I�ɗ�����������������܂��B�ނ�͂��̎����ɓ�����ǂ��Ėk�サ�C�A�t���J���o���ƍl�����܂��B
�uBBC�n���`���v�Ƃ����ԑg�ł́C�ނ�̓X�t�[���ɐ���N�͗��܂�܂������C�Z���ԕX�����I���ĂѕX�����K��C�T�n���₱�̒n�悪�������������ߊg�U���邱�ƂȂ���ł����Ƃ��Ă��܂��B
�������C10���N�O�̐l�ނ̍���T���o�����Ƃ͔��ɓ���̂ŁC�܂����̒n��ł͌������Ă��Ȃ��Ƃ����\���͔ے�ł��܂���B�����Ƃ��C���̂悤�ȏW�c�����Ƃ������Ă������̑��������W�c�ɋz�������ƈ�`�I�ȑ��l���͎����C�o�A�t���J��1�x�Ɍ����邱�ƂɂȂ�܂��B
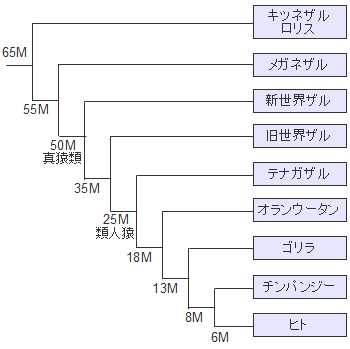
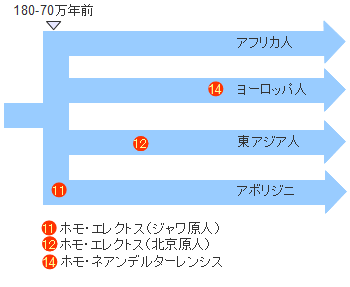
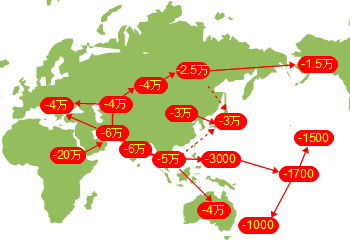
�z���E�T�s�G���X�̏o�A�t���J�͕���������������܂��C���j�ƂȂ鐬���W�c���A�t���J���o���̂�1�x�����ƍl�����܂��B���̏W�c���A�t���J���o��������85,000�N�O���邢��70,000-60,000���N�O�Ƃ���2�̐�������܂��B
����2�̎����̊Ԃɂ̓X�}�g�������g�o�ΎR�̑啬���i74,000�N�O�j�Ƃ������������l�ނɂ��傫�ȉe����^�����C�x���g������܂��B���̒����啬�iVEI=8�j��1991�N�̃s�i�c�{���iVEI=6�j��100�{�ȏ�̋K�͂ł���C�ߋ�100���N�ōő�̕��ł��B�X�}�g�����̓��A���̑唼�͐�ł��C�C���h�ł���10cm�̉ΎR�D���~��ς���܂����B���̉ΎR�D��74,000�N�O�̃}�[�J�[�ƂȂ�܂��B
�C���h�암���W�������v��������ł͒����Ί���g�p�����W�c���g�o�̕��ΑO����ѕ��Ό�����������ĕ�炵�Ă������Ƃ��m�F����Ă��܂��B���̎����ɒ����Ί���g�p���Ă����W�c�̓z���E�T�s�G���X�̉\���������C��������Ɣނ炪�A�t���J���o���̂�74,000�N�O���O�Ƃ������ƂɂȂ�܂��B
8��-6���N�O�̖k�A�t���J�͊��������Ă���C12���N�O�̂悤�ɖk���[�g�Ń��[���V�A�Ɉړ����邱�Ƃ͍l�����Ȃ��ł��B�r�N�g���A����X�[�_���C�G�W�v�g�𗬂ꉺ�锒�i�C��������̎����͂܂��J����Ă��܂���ł����B
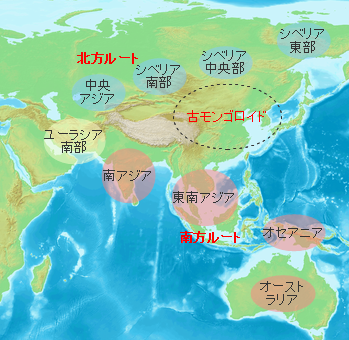
�B��C�l�����郋�[�g�͍g�C�ɂ���Ċu�Ă��Ă���A���r�A�����ł��B�g�C�̂����Ƃ������Ƃ���́u�Q���̖�v�ƌĂ�Ă��镝50km�قǂ̊C���ƂȂ��Ă��܂��B�����͕X���ɂ͊C�ʂ��ቺ��10km�قǂɋ��܂�C���Ȃ����킹���ȒP�Ȕ��̂悤�Ȃ��̂ł��ړ��ł����\��������܂��B�z���E�T�s�G���X�͑�����������C�Y����H���Ƃ��Ă���C�C�ݐ��ɉ����Ĉړ����Ă������Ƃ����l����������o�A�t���J���ƌĂ�Ă��܂��B
����o�A�t���J����⋭����ޗ��Ƃ��Ă̓C�G�����ɂ͐��I�ȐΊ���������Ղ��c����Ă���C�����7���N�O�ɑk��Ƃ��錤��������܂��B�܂��C�A���r�A�����̓암�C�݂ɂ͌��݂͊C��ɒ���ł��܂����^���̕����o���������邱�Ƃ���C�A�t���J���o���W�c�͊C�ݐ��ɉ����Ĉړ��ł����ƍl�����Ă��܂��B�I�}�[�������͂Ƃ���ǂ���ɃI�A�V�X�̓_�݂�����ł����B
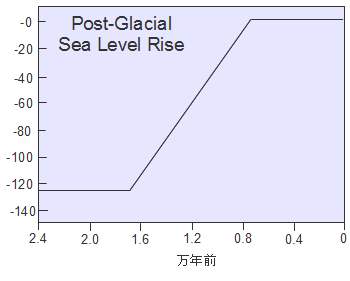
�A�t���J���o���W�c�͊C���ʂ̒ቺ�ɂ��n���̂悤�ɂȂ����z�����Y�C����n��C�����ɓ��B���܂��B����Ńz���E�T�s�G���X�̏W�c�͍����Ɋu�Ă��Ă��Ȃ����[���V�A�암�ɓ��B�����킯�ł��B
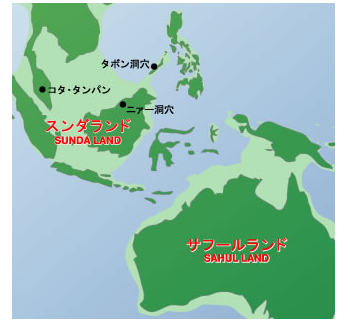
��������W�c�͏��Ȃ��Ƃ�3�ʂ�ɕ�����C��͊C�݉����ɓ��ɁC��͊C�݉����ɐ��ցC������͒����A�W�A�Ɍ������܂����B�������C�����̊���Ŋ��������C��̂��ߐ����[�g�̐�ɂ��郌���@���g�n��i�엀�Ȓ����̎O�����n�сj�͍��������Ă���C1.5���N�قǂ͒x�ꂽ���̂ƍl�����Ă��܂��B
���Ɍ��������W�c�̓C���h�̐��C�݂ł��炭���܂�C���𑝂₵�Ă����܂����B���̌�C���C�݂�쉺���C�r���ŃC���h�����f���C�x���K���p�̉��݂�ʂ�C���悻5���N�O�ɓ���A�W�A�ɓ��B���܂����B
���̉ߒ��ł����Ȃ���ʏW�c�͊��̂悢�y�n�ɗ��܂邱�Ƃ�I�����܂����B���̂悤�ȏW�c�͂���ɕʂ̒n��Ɉړ����邱�Ƃ�����܂������C���ɂ͂����Œ����ԁC�O���Ƃ̌𗬂Ȃ��ɌǗ����ĕ�炵�Ă����W�c������܂��B���̂悤�ȏW�c�ɂ̓A�t���J���o�����̓������悭�c����Ă��܂��B
5���N�O�̓���A�W�A�͌��݂Ƃ͂܂������l�q���قȂ��Ă��܂����B�}���[�����C�X�}�g�����C�{���l�I���C�W���������������ł���C������X���_�����h���`�����Ă��܂����B�I�[�X�g�����A�ƃj���[�M�j�A�����������ł����B�l�ނ̓X���_�����h�ɓ������̂͊m���ł����C�C���h�l�V�A�ɂ����鉻���̓{���l�I���̃j�A���A�i3.5���N�O�̓����ƐΊ�ށj�ȊO�͌������Ă��܂���B
�X���_�����h�����̈ړ����[�g�͂悭�������Ă��܂���B�C���ʂ��������Ă����Ƃ͂����C�`���[�����ƃI�[�X�g�����A�Ƃ̊Ԃɂ�200km�̊C�����݂��Ă��܂����B�ǂ̂悤�ȕ��@�ł�����n�����̂��C�ǂ����Ă��̐�ɃI�[�X�g�����A�����邱�Ƃ����������̂��C�ꕔ�̏W�c�͂��悻4���N�O�ɃI�[�X�g�����A�ɓ��B�i�Y���j���C�A�{���W�j�Ƒ��̂�����Z�����ƂȂ�܂����B�~�g�R���h���ADNA�̕��͂ł��ނ���܂��A�t���J���o���z���E�T�s�G���X�̌����ł��邱�Ƃ��m�F����Ă��܂��B
���[���V�A�암����C�݉����ɐ��Ɍ��������W�c�̓C�����������@���g���A�i�g���A�������C�������̓C�������R�[�J�T�X�������̃��[�g�Ń��[���b�p�ɓ������ƍl�����܂��B�m���ȑ��Ղ��c���Ă���̂̓A�i�g���A�E���[�g�ł��B
�������C�A�t���J�̃T�o���i�N���̃z���E�T�s�G���X�̓��[���b�p�̊���ȋC��ƁC�~�G�̓��Ǝ��Ԃ̕s���ɔY�܂��ꂽ���Ƃł��傤�B�܂��C���[���b�p�S�̂ɍL�����Ă����l�A���f���^�[���l����������Ƃ��ė����ӂ������Ă��܂����B
���[���b�p�ɂ�����ŌẪz���E�T�s�G���X�ł���i�N���}�j�����l�j�̓����̓��[�}�j�A�̓��A�Ŕ������ꂽ���̂ŁC4���N�O�̂��̂Ɣ��肳��܂����B���������Ƃɓ����������Ƃ���A�t���J�n�C�A�W�A�n�C���[���b�p�n�̒��Ԃ̕��e�ł������Ƃ���Ă��܂��B���̐F�͕������Ă��܂���B
����C�l�A���f���^�[���l�̓��[���b�p�ɓK�����������Ɗ��ȑ̋�������C�����ȐΊ���g�p���C���̋Z�p���z���E�T�s�G���X�Ƃ���F�Ȃ������悤�ł��B�C�Y�����H���ɂ��Ă��܂����B�ɂ�������炸�C�z���E�T�s�G���X�Ƃ̐��������ɔs��āC2.5���N�O�ɂ͐�ł��Ă��܂��B
�����z���E�T�s�G���X�ɖ��������̂��͂悭�������Ă��܂��C�z���E�T�s�G���X�̕����Љ�i���W�c�Ԃ̂Ȃ���j�����������̂ŁC�C��ϓ��ȂǂɑΉ��ł����̂ł͂Ƃ�����������܂��B
���[���V�A�암����k�Ɍ��������W�c��4���N�O�ɂ͒����A�W�A����ɂ̓V�x���A�암�ɐi�o���Ă��܂��B2.5���N�O�Ɏn�܂�Ŋ�����ɔ�ׂ�Ƃ��̎����̃V�x���A�암�͉Ă̑����Ɏx����ꂽ��^�����������C�����������Ύ����ɂ���z���E�T�s�G���X�ɂƂ��Ă͏Z�݂悢�Ƃ��낾�����̂�������܂���B
�}�����X�C�уT�C�C�g�i�J�C�Ȃǂ��ނ�̐H���ł���C�g�i�J�C�̖є���܂Ƃ����Ƃɂ��C�����ɑΉ����܂����B�Z���͔��n���Ŋۑ��̍��g�݂Ƀg�i�J�C�Ȃǂ̖є�����������̂��g�p���ꂽ�悤�ł��B���̂悤�ȕ�炵�͌��݂̃V�x���A�����Ńg�i�J�C�ƕ�炷�l�X�̂��̂Ɨގ����Ă����ƍl�����܂��B
�ނ���u�����Ȋw�����فE���{�l�͂邩�ȗ��v�ł��}�����X�n���^�[�Ƃ��ďЉ��Ă��܂��B�}�����X�n���^�[�͊����ɓK�������l�X�ł����B2.5���N�O�̍Ŋ�����ɂ��C���N�[�c�N�s�̖k��80km�ɂ����}���^����Œ�Z�������c��ł��܂����B
��������́C�����̐Ί�ƃ}�����X�̍��Ɖ�C�g�i�J�C�̊p�Ȃǂ��W�ς����e���g���Z���̐Ղ�������������܂����B����48-60�l�C8-10���̉Ƃō\�����ꂽ�W���͉��x�����z�������Ȃ��璷���Ԃ��̒n�ɑ��݂����悤�ł��i���{�l�͂邩�ȗ����j�B
�M�ѐ��܂�̃z���E�T�s�G���X�͊���ȃV�x���A�ŏ��X�ɓK�����Ă����C�k���n�A�W�A�l�ƂȂ�܂����B�����ɓK�p�����ނ�̐g�̓I�����͎��̒ʂ�ł��B
�i1�j �̉��̔��U��h�����߂ɐg�̂͂��肵�葫�͒Z���Ȃ�
�i2�j ��̓�����h�����߂ɕ@�͒Ⴍ�Ȃ蕽�ʓI�Ȋ痧���ɂȂ�
�i3�j �牺���b�����B���āC�܂Ԃ��͌�����d�ɂȂ�
�i4�j �X��̏Ƃ�Ԃ������邽�ߊ�̓A�[�����h�^�ɂȂ�
�i5�j �j���̖̑т������i�Ђ������Ȃ��j
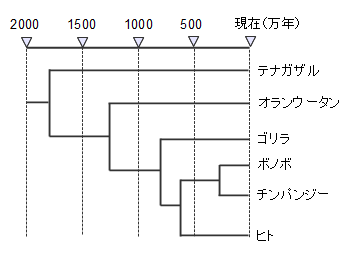
�ނ�͂������ɓ쉺����6000�N�O���ɂ͒����̗D���W�c�ƂȂ��Ă��܂��B����ȑO�̒����ł��Ã����S���C�h���D���W�c�ƂȂ��Ă��܂������C�k���n�A�W�A�l�ɂ����ӕ��ɒǂ�����Ă��܂����ƍl�����Ă��܂��B���̏W�c�̓���ւ��ɂ�蓌�A�W�A�̗D���W�c�ƂȂ����k���n�A�W�A�l���V�����S���C�h�ƌĂт܂��B�V���ɂ͌Â��C�V�����ȊO�̓��ʂȈӖ��͂���܂���B
���͌Ã����S���C�h���ǂ̂悤�ȏW�c�ł����������͂����肵�Ă����Ƃł��B�]���͓���n�A�W�A�l�ł��낤�ƍl�����Ă��܂������C��������Ɠ��{�l�̈�`�q�̊�ɂ͓���n�̂��̂��܂܂�Ă���͂��ł��B
���{�ɐl�ނ��Z�ݎn�߂��̂͂��悻3.5���N�قǑO�̂��Ƃł���C�����̊C�ʐ��ʂ͌��݂�����10m�Ⴍ�C�����C���{�C�������ʏ�ɑ嗤�ƂȂ����Ă���C���{�C�͑傫�Ȍ̂悤�ȏ�Ԃł����B���������āC�����̉ؖk�̗D���W�c�i�Ã����S���C�h�j�����N�������o�R���ē��{�ɂ���Ă��ēꕶ�l�̊�ՂƂȂ����Ƃ����͎̂��R�ȍl�����ł��B
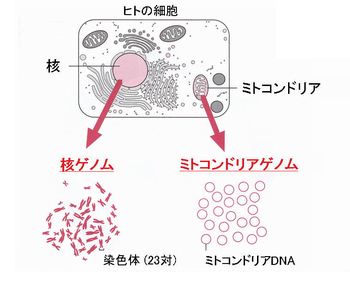
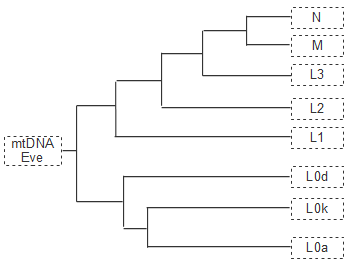
�ꕶ�l�̌��^���l�����ŁC��`�q��͂͗L�͂Ȏ�i�ƂȂ�܂��B������炵���`�B����Ȃ��~�g�R���h���A��`�q�͌����l�ނ̊g�U�̉ߒ��ׂ�̂ɂ悢�w�W�ƂȂ�܂��B����C�������炵���`���Ȃ�Y���F���i�j�����F�́C������XX�C�j����XY�ƂȂ��`�I�Ȑ��ʂ����肳��܂��j���W�c�̗މ��W�ׂ�̂ɓK�������@�ł��B
���݂̓��{�l��Y���F�̂�D2�^��30-70%�i�n��ɂ��قȂ�܂��j�CO�^��0-56%�i�n��ɂ��قȂ�܂��j���߂Ă��܂��BD2�^�͒��N�l�ɂ킸���Ɏc����Ă���ȊO�́C�ǂ��ɂ��Ȃ����������̂ł��BD�^�����W�c�̓`�x�b�g�l(50%)�C�_��̏��������C���i20%�j�C�`�����N�n�A���^�C�i10%�j�CD�c��^�̓A���_�}�������̐l�X�i�ق�100%�j�ƂȂ��Ă��܂��B
�嗤�ɂ�����D�^��Y���F�̂̕��z���l����ƁC���{�l��D�^�͌Ã����S���C�h�R���̂��̂��Ƃ������ƂɂȂ�܂��B�����l�����4���N�O�ɒ����A�W�A���瓌�Ɍ�������D�^�W�c������C�ؖk�̗D���W�c�i�Ã����S���C�h�j�͔ނ�ł͂Ȃ��������Ƃ����l�������o�Ă��܂��B����D�^�W�c�́C���̌�̖k���n�A�W�A�l�iO�^�W�c�j�̓쉺�ɂ����ӂɒǂ����ꂽ��C�z������Ă��܂��܂��B���{�͂��̎����ɂ͌Ǘ��������ƂȂ��Ă�������D�^�����ٓI�Ɏc�����ƍl�����܂��B
���݂̓��{�l��Y���F�̂ɂ�����D2�^�̍�����������l����ƁC�ꕶ�l�̊�ՂƂȂ����̂͑嗤���璩�N�����o�R�œ����Ă���D�^�̏W�c�ł���C����Ɋ����o�R�̖k���n�A�W�A�l�Ɨ����⍕���o�R�̓���n�A�W�A�l�̏W�c��������Ă���ƍl����ƁC�i���Ȃ��Ƃ�Y���F�̂̓��ِ��́j�悭���������܂��B
�ԕX���ɓ�����{�◮�����嗤����Ǘ����邱�Ƃɂ��C�O������V�����l�X���������邱�Ƃ����Ȃ��Ȃ��Ȃ�܂��B�ނ�͋C���g�ŐA�����̐H���Ɍb�܂ꂽ���{�ŋ����E�������C�ꕶ�l���`�����܂����B1���N�ȏ�ɂ킽��Ǘ������W�c�ƂȂ������߁C�ꕶ�l�͔�r�I�C��`�q�̋ψ�ȏW�c�ƂȂ��Ă��܂��B�ꕶ����̓��{�̐l���͉��\�̂悤�ɋC��ϓ��ɂ�葝�����Ă��܂��B�i�S���G�E�l������ǂޓ��{�̗��j�j�B
| ���� |
�l���i���l�j |
�C�� |
| �ꕶ�����i8000�N�O�j
| 2 |
���݂��2���قǒႢ���������� |
| �ꕶ�O���i5500�N�O�j
| 11 |
���݂��1���قǍ����Ȃ� |
| �ꕶ����i3500�N�O�j
| 26 |
�l�����s�[�N�C���≻���n�܂� |
| �ꕶ�ӊ��i3000�N�O�j
| 8 |
�H���s���ɂ��l���͌������� |
| �퐶����i2000�N�O�j
| 59 |
�l�������Ɛ��c���ɂ��l���}��
|
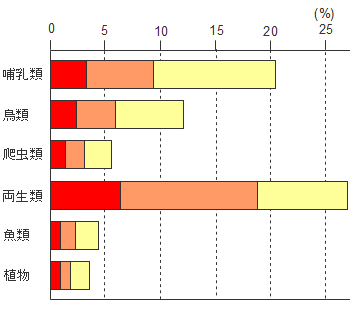
2300�N�O�����琔�S�N�ɂ킽�蒆���嗤�̊e�n���琅�c���Z�p�ƂƂ��ɖk���n�A�W�A�l�̓n���l������Ă��܂��B�n���l�Ɠꕶ�l�͍������C���݂̓��{�l���`������܂��BO�^��Y���F�͓̂n���l�ɂ������炳�ꂽ�ƍl�����܂��B�n���l�ƍ����̏��Ȃ��������邢�͂Ȃ������A�C�k�̐l�����ɂ͂قƂ��O�^�͌����܂���B
���݂̓��{�l�͍������i��ł���C�n��I�ȍ��ق͏������Ȃ��Ă��܂����C��`�q�̃��x���Œ��ׂ�ƗL�ӂ̒n�捷���o�Ă��܂��B�n���l�Ƃ̌��G�����Ȃ��ƍl�����问���̐l�X�ƃA�C�k�̐l�X�̈�`�q�͋߉��x���������Ƃ��������Ă��܂��B�ŋ߂̐l�ފw�̌��_�Ƃ��ẮC����̓��{�l�͖퐶�n��7-8���C�ꕶ�n��2-3���̔䗦�ō������Ă��邻���ł��B