生命の誕生と40億年の進化
生命の誕生(40億年前)
地球は46億年前に太陽系と一緒に形成されました。無数の微惑星を取り込みながら原始地球が誕生します。地球に落下してきた微惑星のもっている巨大な運動エネルギーは衝突の瞬間に大爆発とともに熱に変わります。
地表は微惑星の衝突エネルギーで溶け、マグマの海となりました。微惑星に含まれていた揮発成分は衝突の瞬間に蒸発して水蒸気と二酸化炭素からなる濃密な大気を形成しました。
原始地球の大きさは体積で現在の地球の8分の1程度しかありませんでした。この大きさは軌道上の微惑星を取り込んで成長できる限界です。同様に水星から火星にかけての軌道には原始地球と一緒に成長した20個程度の原始惑星があり、太陽の回りを規則的に周回していました。
しかし、木星や土星といった巨大ガス惑星が成長し、その引力影響により原始惑星の軌道は乱され、他の原始惑星の軌道と重なるようになります。原始金星や原始地球は周辺軌道の原始脇制と次々に衝突・合体(ジャイアントインパクト)を繰り返し成長していきます。
金星は6-8個、地球は7-9個程度の原始惑星が合体したと考えられています。ジャイアント・インパクトの度に原始地球はマグマの海となり、公転面に対する地軸の傾きも変化しました。
最後のジャイアントインパクトにおいて原始地球に衝突した原始惑星は自分が砕けるとともに地球のマントルの一部をはぎとり、無数の破片が地球の周囲を回るようになります。
衝突のエネルギーにより地球はマグマの海となり、その周辺を高温の破片が周回します。破片の一部は地球の重力により落下し、残ったものは一つのまとまりとなり、私たちが月と呼んでいる天体となります。
月は原始惑星の一部とと地球のマントルの一部が融合したものですので、地球の岩石と組成がよく似ています。月を生み出したジャイアントインパクトは地球にとっては最後のものとなります。
最後のジャイアントインパクトの結果、原始地球は地球となり、現在よりもはるか近いところに月が周回しています。太陽系の誕生は46億年前ということになりますが、地球の誕生は最後のジャイアントインパクトとするのが天文学の主流のようであり、その考えでは地球も月もおよそ45億5000万年前に誕生したとされています。
最後のジャイアントインパクトにより、地球の公転面に対する自転軸の傾きを現在の約23.4度になります。これはその後の地球環境において決定的に重要な役割を果たします。また、すぐ近くを大きな天体が周回することにより、地軸の傾きのブレが最小限に抑えられるようになりました。
ジャイアントインパクト直後の月は2万㎞ほどのところを周回し、地球の自転速度は現在の3-4倍であり、1日は6-8時間程度でした。しかし、月は地球の潮汐力により少しずつ角速度を得て地球から遠ざかり、同時に地球の角運動量は減少していきます。現在の月は地球から38万㎞のかなたにあり、地球の1日は24時間となっています。
地球誕生後、時間とともに微惑星の衝突も減り、地表温度も徐々に下がり出します。地表のマグマは固化し薄い岩石の皮膜を形成します。
それでも地表温度は数100℃もあり、水蒸気と二酸化炭素からなる200気圧、数100kmに達する濃密な原始大気の中では激しい対流が起きます。原始大気の表層では冷やされた雲が雨になります。しかし、雨はまだ熱い地表付近で熱せられ再び水蒸気に戻ります。
地表の温度がさらに下がると、ついに原始大気中の水蒸気が生み出した雨が地表に到達します。地球史上で最大の豪雨だったことでしょう。地表に到達した雨は蒸発しますが、地表の熱を効率よく宇宙空間に放出していきます。
何回かの蒸発サイクルにより海が誕生します。海の誕生時期についてはいくつかの学説がありますが43億年前というのが有力です。海の誕生は地球の進化、生命の誕生において決定的に重要な役割を果たします。
しかし、その後も地球レベルの破局的な衝突は続きます。現在の月の表面は誕生後の隕石の衝突により形成された無数のクレーターに覆われています。月のすぐ近くにあり、ずっと大きな地球には(重力も大きいので)最後のジャイアントインパクト後も月の何倍もの微惑星や隕石が降り注いだはずです。
確率的には直径が100-500kmもの微惑星が複数回衝突したと推計されます。このような巨大な天体が衝突すると落下地点の岩石は瞬時に蒸発し、高温の岩石蒸気が地球の全表面を覆うことになります。そのため海は沸騰し完全に干上がります。このようなイベントは生まれたばかりの生命にとっては致命的なものになります。生命は生成と消滅のサイクルを何回か繰り返したかもしれません。
現在の学説では地球が誕生してから6億年ほど経った頃(40億年前)、海で生命が誕生したといわれています。当時の地表は強い紫外線や荷電粒子が容赦なく降り注ぎ、生命にとっては致命的な環境でした。生命が存在できる環境は海中だけでした。
原始の海には生命に必要な有機分子(アミノ酸、核酸塩基、糖、脂肪酸、炭化水素など)が豊富に存在していたと考えられています。それは、星間物質に含まれ小天体と一緒に地球に到達したものもあれば、紫外線、荷電粒子、落雷などにより活性化されていた地球の原始大気中でできたものかもしれません。
生命の材料を星間分子雲に求める学説もあります。地球生物が作り出すアミノ酸の光学異性体型がほぼすべて左手型であり、宇宙の星間分子雲に存在するアミノ酸も(右手型アミノ酸は宇宙線により破壊されやすいことから)左手型となります。このため、原始生命のもとになったアミノ酸は宇宙起源のものであるとしています。
日本の小惑星探査機はやぶさ2に大きな期待がかけられているのは、小惑星「りゅうぐう」には太陽系形成時の始源物質が変成されない形で保存されているからです。地球の水の起源ですら、岩石型の小惑星か、その後に加わった氷型のすい星かの議論は決着していません。生命に必要な基本物質は小惑星から見つかれば、生命材料の宇宙起源が有力な仮説となります。
起源はどうあれ生命の材料に溢れていた原始の海で生命は誕生しました。海水に溶けこんだ有機物が化学進化により最初期の生命である原始生命体になったという化学進化説が現在の主流となっています。
化学進化はアミノ酸や核酸が化学的にくっついたり離れたりしている中から、しだいにたんぱく質と核酸を薄い膜の中に収め、自己の形をもち、増殖することが出来るようになったと考えられています。生命は身近にある材料を組み合わせて誕生し、その基本構成は現在まで受け継がれています。
しかし、この化学進化が生命誕生の唯一の考え方というわけではありません。生命とは神の御手によって創造されたものとする考え方や、生命の起源は地球外にあるという考え方も併存しています。
化学進化の一つの考え方に表面代謝説があります。それによると、『黄鉄鉱(FeS2)表面で有機物の重合反応を含めたあらゆる化学反応が発生した、初期の生命は単位膜によって覆われず黄鉄鉱表面に存在する代謝系であった』となっています。
単位膜がないことや自己複製能力がないことから、生命と定義するのは無理があると思いますが、化学進化がこのような環境下で進んでいく可能性があるという一つの方向を示しています。
現在、初期の生命(生物)に関しては原始生命体と共通祖先(LUCA)という概念があります。時系列的には化学進化→原始生命体→共通祖先ということになりますが、原始生命体は化学進化のどこで生命の線引きをするかという概念であり、共通祖先は生物を遺伝子解析から系統づけた結果として生まれた概念です。両者は時間的なギャップと同時に概念の相違があります。生命は原始生命体に始まりますが、生物進化は共通祖先から始まるということです。
生命の定義(生命としての要件)には次の3点があげられます。
(1)代謝系を有する
(2)細胞という形状を有する
(3)自己複製が可能である
しかし、「表面代謝説」にもあるように初期生命の定義があいまいになりつつあります。また、さなざまな化学進化モデルも提案されています。その意味では「原始生命体」をどのように定義するかは、生命誕生を論じるうえではとても重要になっています。代謝系と栄養の面から原始生命体がどのようなものであったか考えてみましょう。
生物の代謝系は異化代謝(大きな分子を小さな分子に分解する)と同化代謝(小さな分子から大きな分子を合成する)に分かれます。代謝の基本的なものはエネルギー代謝であり、異化代謝がより早い時期に成立したと考えられています。
| 代謝の分類 | 説明 | 成立時期 |
|---|---|---|
| 化学合成 | 化学物質を利用した同化代謝 | もっとも早い時期に成立 |
| 嫌気呼吸 | 酸素を利用しない異化代謝 | もっとも早い時期に成立 |
| 好気性呼吸 | 酸素を利用する異化代謝 | およそ35億年前に成立 |
| 光合成 | 光を利用した同化代謝 | およそ30億年前に成立 |
嫌気呼吸とは酸素を用いない異化代謝系の総称です。この分類の中に含まれる嫌気的解糖は発酵と呼ばれ、すべての生物が備えているので、もっとも古い時期に成立した代謝系であると考えられます。
原始生命体は炭素源として何を利用していたのかという点も重要な論点です。化学進化を提唱したオパーリンは原始生命体は原始海洋中に既に存在していた有機物を代謝する従属栄養(炭素源を有機物に依存している)生物であったとしています。つまり、周辺の有機物を取込み、それを代謝していたということです。
それに対して、最近は原始生命体は独立栄養(二酸化炭素から有機物を同化代謝できる)的であったとする学説が有力になっています。それは、1970年代に中央海嶺の熱水噴出孔で化学合成による独立栄養細菌群が発見されたことによります。
このような化学合成細菌は熱水中の硫化水素のような還元的な無機物を酸化することによりエネルギーを得て、二酸化炭素から糖を同化代謝することができます。
一方、生物進化の観点から見ると系統樹では根が深い生物群はそのほとんどが化学合成独立栄養的に生育し、好熱性を示すという結果が出ています。このことは、共通祖先が熱水噴出孔周辺の化学合成細菌に近いものであることを示唆しています。
チムニーと呼ばれる熱水噴出孔から煙突のように伸びる柱状の構造物は、熱水に溶けていた物質が冷たい海水に冷やされて抽出されたものであり、多孔質の構造をもっています。
このような構造は生命の材料を濃集し、化学進化にとっては都合のよい環境でもあります。生命が誕生したころの海底は至る所で熱水噴出孔のような構造があったと考えられています。
太陽光のまったく届かない熱水噴出孔で光合成生物への準備が行われているという報告もあります。その細菌は熱水噴出口からの赤外線を感知して移動することにより、熱水噴出口の周辺に集まることができると考えられています。
赤外線では光合成を行うには不十分ですが、光を検出する仕組みが光合成細菌への進化につながったのではという考え方です。実際に太平洋の熱水噴出口から光合成細菌と思われる細菌が単離されたという報告もあります。
このページの目次
- 生命の誕生(40億年前)
- 生物共通祖先と系統樹の考え方
- シアノバクテリアの出現(30億年前)
- 猛毒の酸素が生物の進化を促しました
- 磁気圏の形成(27億年前)
- 鉄の酸化と酸素の増加(27-20億年前)
- 多細胞生物の出現(10億年前)
- カンブリア爆発(5.5億年前)
- オゾン層の形成(5億年前)
- 生物の大繁栄(5.1-2.45億年前)
- 恐竜の大繁栄(2億年前)
- 哺乳類の時代(6500万年前~)
- 大絶滅は何回か発生しました
| 項目 | 質量(10億トン) |
|---|---|
| 地球 | 6 X 1012 |
| 地球表面の水 | 1.4 X 109 |
| 海水 | 1.35 X 109 |
| 氷床 | 2.5 X 107 |
| 1年間の海水蒸発量 | 500,000 |
| 1年間の陸上降水量 | 50,000 |
| 大気中の酸素 | 1,100,000 |
| 大気中の二酸化炭素 | 2,750 |
| 大気中の炭素 | 750 |
| 陸上生物中の炭素 | 610 |
| 陸上植物の年間炭素固定量 | 120 |
| 土壌中の炭素 | 1600 |
| 大陸の化石炭素 | 4,000 |
| 海洋表層の無機炭素 | 1,000 |
| 海洋生物中の炭素 | ■ |
| 海洋生物の年間炭素固定量 | ■ |
| 深海に蓄積されている炭素 | 38,000 |
| 海水中の塩分(NaCl) | 40,000,000 |
| 海水中のマグネシウム | 2,000,000 |
| 海水中の金 | 0.006 |
現在の地球表層の物質、40億年前に誕生した生命は地球表層の物質循環に大きな影響を与えました。
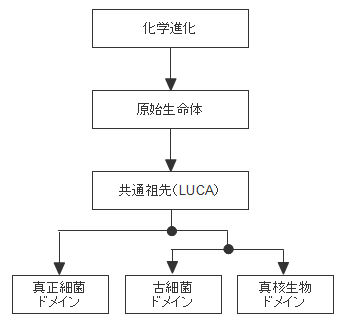
生命進化の概念図、原始生命体とは化学進化の結果に生まれた生命であり、共通祖先が誕生する以前の生命と定義されています。共通祖先とは真正細菌と古細菌の共通の祖先という意味であり、生物進化上の定義ということになります。生命は原始生命体に始まりますが、生物進化は共通祖先から始まるものということです。
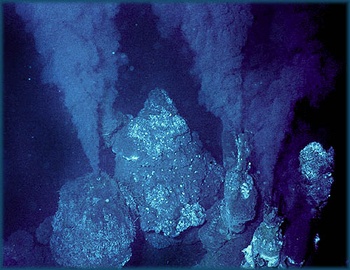
中央海嶺リフト・バレーのブラックスモーカー、画像は「amnh.org」から引用しました。チムニーと呼ばれる熱水析出物からできた塔の上部から黒い煙のような熱水が噴き出しています。ここには化学合成細菌を一次生産者とする濃密な生態系があり、生命誕生の場ではないかと考えられています。

化学合成細菌がエネルギーを産生する仕組み、化学合成菌は現在でも嫌気環境で生息しており還元的な物質を酸化することにより化学エネルギーを産生し、それを利用して二酸化炭素から有機物を合成します。初期の生物は好熱型であり、独立栄養であったと考えられています。
生物共通祖先と系統樹の考え方
現在(過去に存在した)の生物の系統樹を作成するうえでもっとも困難であったことは真核生物(遺伝子を収める核をもつ生物)と原核生物(核をもたない生物)の関係でした。
生物進化には「単純な生物から複雑な生物へ進化する」という大原則があります。それに基づくと単純な原核生物から複雑な真核生物に進化することになりますが、両者は非常に異なっており、単純な進化から説明がつきませんでした。
その後、古細菌が発見され、原核生物と真核生物の分岐の前に古細菌と真正細菌の分岐が起きていることが分かりました。古細菌から真核生物が分岐したという考え方は、広く受け入れられるようになり、これにより、1990年には地球上の生物は共通祖先から進化していったという3ドメイン系統樹を完成させることができました。
ドメインとは生物の最上位の分類であり、真正細菌、古細菌、真核生物の3つのドメインからなります。地球上の生物はすべてこの3つのドメインのいずれかに属することになります。この系統樹の根に相当するものが共通祖先ということになります。ただし、生物の基本的な分類は時間とともに大きく変化してきました。
1735年にスウェーデンの博物学者、生物学者、植物学者であるリンネ(1707-1778年)は「自然の体系」の中でそれまでに知られていた動植物についての情報を整理して分類表を作り、生物分類を体系化しました。また、彼は生物種の学名を属名と種小名からなる2語のラテン語で表す体系化した命名法を作りました。
現在の生物種の学名はリンネの命名法を踏襲し、国際的な命名規約に基づいて決定されています。リンネは種の特徴と類似する生物との類似点・差異点を明らかにし、属、目、綱、門、界といった上位の階層的な分類単位を制定しました。現在の生物分類学の基礎を作り上げた業績により「分類学の父」と呼ばれています。
リンネは生涯現役の学者であり、世界中から膨大な標本や資料を収集しました。彼の死後、リンネの妻は夫の遺言にしたがい、蔵書や原稿、収集品のほとんどをしっかりと管理してくれそうな人物(ジェームズ・スミス)に売却しました。スミスは1788年に貴重な品々の受け皿となる学会を設立しロンドン・リンネ協会と名づけました。
ロンドン・リンネ協会は世界でもっとも古い学術組織であり、ここにはリンネが世界中から収集した19,000もの生物標本が保管されています。現在は研究目的に限定して公開されていますが、まもなくデジタル化の完成により一般公開されるかもしれません。リンネは知識は伝達され、使われるためにこそあるという信念をもっていましたので、デジタル・コンテンツの公開をきっと喜んでくれることでしょう。(National Geographics の記事より)
リンネの生物分類は「植物」と「動物」の2界となっていました。無理もありません、ダーウィンが「種の起源」を発表したのは1858年のことです。そういえば、ダーウィンもロンドン・リンネ協会の会員でした。
17世紀に微生物が発見(見えるようになった)されましたが、この微生物が目に見える動物や植物とどのような関係があるかはまだ理解されていなかったのです。微生物が生物の分類に含まれるようになったのは1894年のことです。
1977年に古細菌が生物分類の中に入り、6界説となりましたが、1990年には3ドメインに整理されました。現在の真核生物ドメインは原生生物、菌類、植物、動物を含みます。しかし、生物の分類は遺伝子解析から新たな展開を見せており、動物と菌類は同一の系統に含まれるとして上位分類を設けるという考え方もあります。
生物の系統分類をするうえでリンネのとった方法は外観、解剖など目に見える違いによるものでした。現在の系統分類では形態、発生などの要素が加算され、最近では遺伝子がその重要な手がかかりとなっています。
遺伝子分析では二つの種について特定の遺伝子を調べ、塩基配列が何ヶ所違っているかを数値化します。この数値により二つの種がどのくらい近い(遠い)関係にあるかが分かります。また、近縁の種であれば、二つの種が共通の祖先から分岐してどのくらいの時間が経過しているかを表しています。
この考え方を分子時計といいます。分子時計という考え方は遺伝子の塩基配列の変化は突然変異により起こり、その発生確率は一定であるという前提に基づいています。一つの塩基配列が変化する確率時間(t)が分かれば、(n)個の変異が起きるのに必要な時間は(nt)ということになります。
この分子時計を使用して類人猿の中でチンパンジーが分岐したのは1250万年前、ゴリラが分岐したのは800万年前、チンパンジーとヒトが分岐したのは600万年前というように経過時間を含めた系統樹を作ることができます。この分子時計は理論値であり、実際の系統樹には化石などの具体的な証拠が必要です。
1986年にカール・ウーズによって示された3超界(3ドメイン)分類も、すべての生物がもっているタンパク質合成系の共通要素の1つである、リボソームRNA遺伝子の解析から得られたものです。
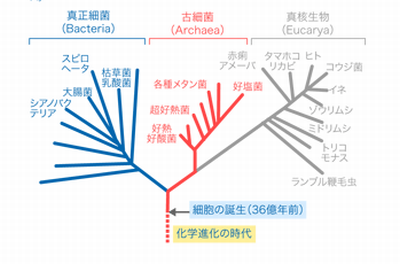
生物の系統樹の一つの形、画像は「実験医学online」から引用しました。生物進化の観点から見ると系統樹では根が深い生物群はそのほとんどが化学合成独立栄養的に生育し、好熱性を示すという結果が出ています。つまり、共通祖先はそのようなものであったということになります。
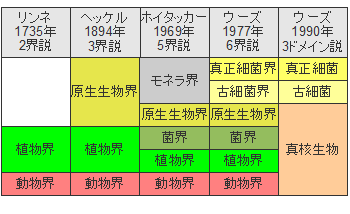
生物分類の変遷、界より上の階級はsuperkingdom、empire、domain などと呼ばれていましたが、現在ではドメインで統一されています。分類学の父と呼ばれるリンネは動物と植物を対象にしましたが、現在では生物全体を真核生物、真正細菌、古細菌という3つのドメインに分類しています。

分類学の父と称されるリンネの肖像、画像はwikipedia から引用しました。彼のラテン語による二命名法は現在でもそのまま使用されています。
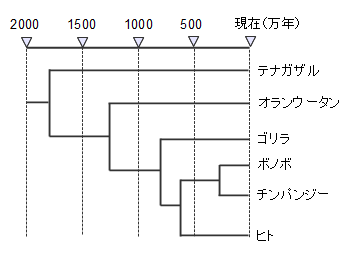
分子時計を用いた類人猿の系統樹、遺伝子の変化速度が一定とすれば、遺伝子の違いから共通の祖先から分岐してからの経過時間が分かります。
シアノバクテリアの出現(30億年前)
共通祖先から細菌と古細菌が分岐してからの数億年は、両者とも食料とエネルギーという制約を受けながら、命をつないできました。彼らの中で従属栄養型の生物は自分の体を作ったり、生命活動に必要なエネルギーを外部から得る必要があります。しかし、彼らの生存していた環境に存在する化学的に合成された有機物は彼らが大繁栄するほど多くはなかったことでしょう。
化学合成細菌のように独立栄養型の生物でも二酸化炭素から有機物を作るためには、硫化水素のように還元的な物質を酸化してエネルギーを得る必要があります。しかし、そのような物質も海底の一部に限定されていたことでしょう。
この古い世界の中にも小さな変化が起きました。光をエネルギーとして利用する光合成細菌が出現したのです。光合成を行う細菌にはシアノバクテリア、紅色細菌、緑色硫黄細菌、緑色非硫黄細菌、ヘリオバクテリアなどがあります。
遺伝子解析(16S_rRNA系統解析)によると光合成細菌というまとまりはなく、上記の細菌類はそれぞれ別の系統に属しています。光をエネルギーとして利用する細菌は異なった代謝システムをもっているということになります。
二酸化炭素から有機物を作るためには水素が必要です。この水素をどのように得るかにより、代謝システムが異なります。例えば緑色硫黄細菌は硫化水素(H2S)から水素を取り出すため、廃棄物は硫黄ということになります。この仕組みはある特定の物質を必要としているという観点からは、化学合成型の細菌と類似しています。
これに対しておよそ30億年前に出現したシアノバクテリアは水素の供与体として水を選択したため、海の広い範囲で繁栄する可能性が広がりました。シアノバクテリアは水素の供与体に水を選んだので、光合成の廃棄物は酸素ということになります。
水、二酸化炭素という海水中に豊富にある物質を利用し、光のエネルギーで有機物(糖)を作りだすというシアノバクテリアの選択は大きな成功を収めることになります。その結果、彼らの放出する遊離酸素が海水中に増加することになります。酸素は物質を酸化するため、低酸素環境で暮らしてきた生物にとっては猛毒物質でした。
しかし、シアノバクテリアはとその他の細菌は排他的な関係とはならなかったようです。現在のバクテリアマット(バクテリアがマット状に積み重なった生態系)でも両者が共存している様子が観察されます。
河口のデルタに見られるバクテリアマットの場合、最上部は緑色のシアノバクテリア、中間層には紅色光合成細菌、最下層が従属栄養型の嫌気性バクテリアというように、垂直方向での棲み分けができています。紅色光合成細菌はシアノバクテリアが使わない波長成分を利用して、光合成を行っており、光をシェアしています。
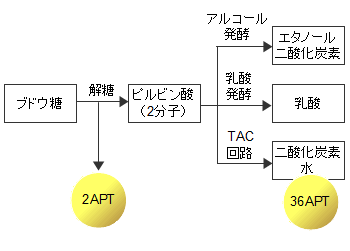
酸素の少ない環境では微生物は発酵により細々とエネルギーを調達します。アルコール発酵では2個のATPしか得られませんが、クエン酸回路による有酸素燃焼では平均して38個のATPを得ることができます。この仕組みはほとんどの生物に共通しています。
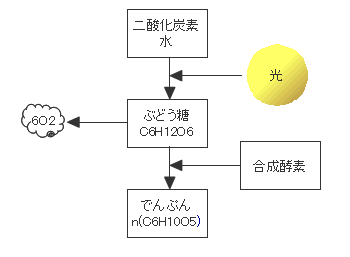
光合成(水と二酸化炭素からブドウ糖を産生する)生物は30億年前に現れました。光合成生物の放出する酸素により地球環境は大きく変化しました。地球上の生物の大半は光合成から始まる食物連鎖により生きています。
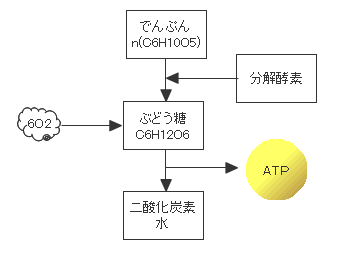
生物がでんぷんを分解してブドウ糖を産生し燃焼させる仕組みはほとんどの生物に共通しています。30億年前に始まった光合成は、生物にとって地球でもっともすぐれたエネルギー生産の仕組みであったようです。
猛毒の酸素が生物の進化を促しました
シアノバクテリアの放出する酸素は猛毒物質でしたが、酸素をうまく利用することができれば酸素呼吸が可能となり、いままでの嫌気呼吸に比べると20倍近いエネルギーを産生することができます。この環境の変化がそれまでの生物に進化を促します。
ある種の従属栄養型の細菌類は酸素と共生する道を選び、酸素呼吸により取り込んだ有機物を代謝して、大きなエネルギーを利用する能力をもち、好気性バクテリアとなりました。このエネルギー革命により大きな運動能力も獲得し、肉食性のバクテリアとなったようです。捕食、被捕食という新しい関係が生まれました。
古細菌のグループはほとんど猛毒の酸素から逃れ、酸素の少ない環境で暮らすことを選択しました。しかし、古細菌の中で細胞壁を失い、柔らかい細胞膜だけをもったものは海底に残ることを選択しました。
一般的に原核生物は固い細胞壁をもっています。細胞壁は細胞の構造を支え、物理的・化学的な刺激に対する防御の役割をもっていました。しかし、古細菌の一部には、周辺の有機物が少なくなってくると細胞壁を捨てて、柔らかい細胞膜でくるまれた形態をとるものが現れました。柔らかい膜をもつ細胞は効率的に有機物を取り込むことができたと考えられています。
しかし、肉食性の好気性バクテリアが現れると、細胞壁を捨てた古細菌は生存の危機に立たされます。肉食性のバクテリアに対抗するため、彼らは複数の仲間と合体して細胞を大きくする道を選択しました。
古細菌がもっていた遺伝子は一つにまとめられて、その外側に膜が作られました。これが核といわれる構造です。こうして、核をもち、柔らかい細胞膜でくるまれた大きな細胞が誕生しました。これが、これが原始的な真核生物のはじまりと考えられています。
原始的真核生物は進化を続け、20億年前には真核生物が登場します。とはいうものの、従属的栄養型で酸素呼吸をもたない原始的真核生物は大きく繁栄することはできませんでした。
現在の真核生物で大きなグループを形成しているのは原生生物とよばれる単細胞生物です。原生生物は細胞の微細構造で特徴づけられる数十の群に分類されます。近年の分類学の成果により、微細構造の違いは異なる起源によるものであることが明らかになっています。
しかも、遺伝子分析による系統樹ではこのような多数の系統群がほぼ同じ時期(10-12億年前)に分岐していることも明らかになっています。真核生物にとって何か重大な変化がこの時期あるいはこの時期以前にあったことがうかがえます。その変化とは原始的真核生物と真正細菌の合体あるいは細胞内共生であったろうと考えられます。
現在の真核生物の細胞はミトコンドリアと葉緑体という細胞内器官をもっています。葉緑体は植物が光合成を行うときの主役となるもので、その機能はシアノバクテリアと類似しています。ミトコンドリアは細胞内でエネルギーを産生する機能をもっており、その機能は好気性バクテリアと同じです。
真核生物の細胞においてはすべての遺伝子は核に収められているはずですが、葉緑体とミトコンドリアは核の外に独自の遺伝子をもっており、それらに由来する遺伝を細胞質遺伝といいます。つまり、真核生物の細胞に対して葉緑体やミトコンドリアは半分独立した存在だということができます。
このことから、なんらかの原因で大きな細胞をもつ真核生物の中にシアノバクテリアや好気性バクテリアが入り込んだ(取り込まれた)と考えられています。現在主流となっている細胞共生説では、真核生物の細胞はそのようなバクテリアに安定した環境を提供する代わりに、彼らの産生する有機物やエネルギーを利用する共生関係が形成されたとしています。
長い共生関係の間にシアノバクテリアや好気性バクテリアの遺伝子の一部は真核生物の核に取り込まれましたが、一部は核外に残されているということになります。
このような共生関係がどのようにして生じたかについては結論が出ていません。原始真核生物が好気性バクテリアを捕食したが、消化しないで自分の細胞内に住まわせるようになった(捕食説)、メタン生成古細菌と好気性細菌が互いに廃棄物をやり取りして共生するようになった(水素説)などいろいろな考えが提案されています。
現在の生物においても共生はごくあたりまえのことです。もちろん、最初から何かを補完しあう関係であったというより、一方的な寄生状態から始まった関係が多いと考えられます。
寄生生物が宿主から一方的に搾取する関係を続けていると、宿主の方にもそれに対する防御反応が発生します。また、あまりにも宿主に過大な負担をかけると宿主の死を招いてしまうことにもなります。つまり、一方的な寄生は長続きしないことになり、時間とともに共生関係に移行していくものと考えられます。
生物間の共生については興味深い実験結果(Jeon and Lorch、1967)が報告されています。この実験はアメーバを使用するものでしたが、不注意から未知の複数の細菌(X-細菌)に感染させてしまい、その毒性によってアメーバを死なせてしまいました。
ところが、生き残ったアメーバの中には感染前の増殖能力を回復したものが現れ、 興味深いことにそれらのアメーバの中では X-細菌も一定数を維持しつつ、アメーバとともに増殖していました。
さらに驚くべきことは, このようにして共生関係が成立したアメーバとX-細菌はもとは単独で生活できていたにもかかわらず、5年後に分離すると、どちらも単独では生きられず死んでしまいました。これは生物間の共生が生物の形質を変化させることが実験室で初めて観察された事例となりました。
敵対する生物と短時間で共生関係を構築できるというこうことは、細胞レベルにおける共生の可能性が私たちが考えているよりずっと複雑、かつ大きいことを示しています。
真核生物の遺伝子は古細菌由来のものと真正細菌由来のものが交雑しており、それが真核生物の多様化をもたらした一つの要因となっているとも考えられます。ともあれ、ミトコンドリアやシアノバクテリアという酸素時代の新しい生物を取り込むことにより、真核生物は酸素という新しい環境で急速な繁栄を開始することになります。初期の真核生物の他の系統は新しい真核生物の爆発的な繁栄の陰で絶滅したものと考えられます。
生物進化の原動力となったシアノバクテリアの直接の子孫は現在でもオーストラリア西海岸のハメリンプールと呼ばれる浅い海で生息しています。写真のようにほとんど岩のように見えるものはストロマトライトと呼ばれており、シアノバクテリアが作り出した構造物です。内部は層状になっており、毎日少しずつ付着した砂を粘液で固定しながら成長しています。
そのため、成長したストロマトライトはマッシュルームのように上にいくほど大きくなります。この海は乾燥地帯に位置し、かつ外海との海水の出入りが少ないので、塩分濃度が非常に濃くなります。このような環境では他の生物との競合が少ないので古い生物が生息できるようです。
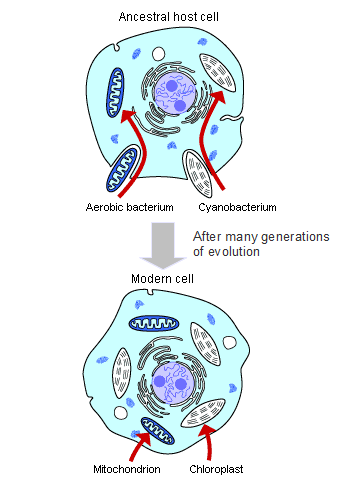
共生説による真核生物の誕生、画像は「Understanding evolution」より引用し一部変更しました。
Ancestral host cell: 受け入れ先となった祖先の細胞
Aerobic bacterium: 好気性バクテリア
Cyanobacterium: シアノバクテリア
Mitochondrion: ミトコンドリア
Chloroplast: 葉緑体
共生説では大きな祖先細胞(原始真核生物)に酸素呼吸の能力を獲得した好気性バクテリアや光合成の能力を獲得したシアノバクテリアが共生して真核生物に進化していったとされています。

オーストラリア西海岸のシャークベイに現在も生き続けているストロマトライト、写真は「ストロマトライト」から引用しました。塩分濃度が高く、他の生物との競合が少ない環境で、シアノバクテリアが作る岩のように見えるストロマトライトが多数あります。ストロマトライトが大繁栄した時期の海岸はこのような光景であったのかもしれません。
磁気圏の形成(27億年前)
27億年前に地球は磁気をもつようになりました。例えて言うなら地球の北極と南極に合わせて(多少はずれていますが)巨大な棒磁石を置いたようなものです。現在は北極の近くにS極があります。
この巨大な磁石は鉄やニッケルでできている地球の外核がゆっくりと対流しているためだと考えられています。高校の物理で電磁誘導について学習したことと思います。磁場の中でコイルを動かすとコイルに電流が流れるといういう現象です。
動かすものはコイルでなく金属であっても電流は発生します。地球の内核は固体ですが、外核は液体の金属ですので対流を起こします。磁場があれば対流により電流が流れ、その向きが元の磁場と同じであれば、地球は磁場を維持することができます。これは自己励起型発電機と同じ原理です。
したがって、きっかけの初期励起があれば地球は磁場をもつことになります。当然、地球の内部には磁気に見合う大きな電流が流れています。この電流を流すエネルギーはどこからきているかというと、それは外核の対流からということになります。水力発電では水の位置エネルギーで発電機を回すように、地球は熱的な対流で発電機を回しているのです。
地球が磁気をもつことにより、地球の外側に磁気圏が形成されます。この磁気圏が太陽からの太陽風のバリアとして働いてくれます。核融合により巨大なエネルギーを生み出している太陽からは絶え間なく太陽風が噴き出してきます。太陽風はほとんどが水素なのですが、高温のため電子と陽子に分離したプラズマ状態になっています。
プラズマとは気体を構成する分子が電子と陽イオンに電離した状態で自由に運動している状態です。プラズマ中の電荷は全体として電気的に中性な状態に保たれますが、個々の構成要素は荷電粒子ということになります。水素ガスの場合は陽イオンはそのまま陽子ということになります。
プラズマのような荷電粒子は磁場を横切ることができませんので、地球の磁場の勢力圏に入ることができず、磁場を押しつぶすようにしてその縁を移動ることになります。つまり、磁気圏は太陽風から地球を守るバリアとなっているわけです。
高エネルギーの荷電粒子は地球上の生物には非常に危険です。地球ができてから磁気圏ができるまでのおよそ30億年間は地表には宇宙から大量の高エネルギー荷電粒子が降り注いでいました。さらに、太陽光に含まれる波長の短い紫外線、高エネルギーの宇宙線なども生物にとっては脅威となります。
磁気圏ができることにより光合成を行うシアノバクテリアはより安全に、より活発に増殖できるようになったことでしょう。実際、この時期からシアノバクテリアは大繁殖するようになります。
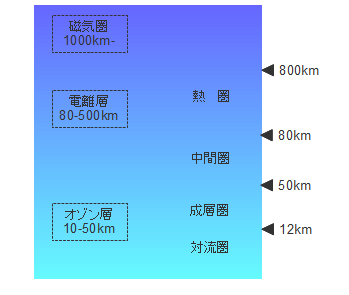
地球の大気、地球の大気圏は対流圏、成層圏、中間圏、熱圏に区分されます。熱圏の上に外気圏をおく場合もあります。オゾン層は太陽光に含まれる生物にとって有害な波長の短い紫外線を吸収するバリアとなっています。磁気圏、電離層は太陽風や宇宙線のような高エネルギーの荷電粒子に対するバリアとなっています。生命が誕生した頃の海も重要なバリアとして機能していました。現在の地球は何層ものバリアに守られ、生物にとっては安全な環境となっています。
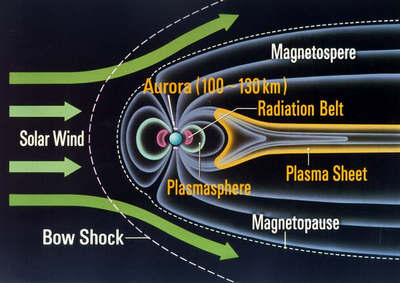
太陽風を遮る地球磁気圏、画像は「宇宙科学研究所」のものを引用しました。太陽風とは太陽から吹き出してくるプラズマ(高温のため電子と原子核が電離した状態の気体、水素の場合は電子と陽子が分離した状態)であり、その主体は水素です。太陽風は磁場を横切ることができませんので、地球の磁場の勢力圏に入ることができません。地球磁気圏のおかげで生物は有害な高エネルギー荷電粒子から守られています。この地球磁気ができたのが27億年前ということになります。
鉄の酸化と酸素の増加(27-20億年前)
シアノバクテリアの大繁殖により海水中に放出された酸素は海水中に溶け込んでいた鉄を酸化するのに使用されました。酸素の少ない環境では安定して水に溶けていた鉄イオンは酸化により不溶性のものになり、水酸化鉄(Ⅲ)Fe(OH)3として海水から析出して沈殿しました。この水酸化鉄が脱水すると赤鉄鉱Fe2O3となります。
 水酸化鉄(Ⅲ)も赤鉄鉱と同じように赤褐色の物質です。このような水酸化鉄の沈殿はシアノバクテリアが大繁殖していた浅い海で起こりました。当時の海岸線は鉄さび色に染まったことでしょう。
水酸化鉄(Ⅲ)も赤鉄鉱と同じように赤褐色の物質です。このような水酸化鉄の沈殿はシアノバクテリアが大繁殖していた浅い海で起こりました。当時の海岸線は鉄さび色に染まったことでしょう。
このとき堆積した水酸化鉄から縞状鉄鉱床が形成されました。この鉱床は数十mからときには数百mにもなります。鉱床は鉄鉱石に富む赤茶色の部分と主にケイ酸塩鉱物からなる薄茶色の部分がそれぞれ厚さ5-30mmほどの互層になっており、断面が縞状に見えることから縞状鉄鉱床と名づけられました。
 縞状鉄鉱床の大部分は27-20億年前に形成されたものですが、7億年前にも大規模な縞状鉄鉱床が形成されました。縞状鉄鉱床ができるためには無酸素の海中に鉄イオンが溶け込むことが必要です。すでに酸素の十分にあった時期にどうして縞状鉄鉱床が生成されたかについては、大氷河期(スノーボール・アース)との関連が指摘されています。
縞状鉄鉱床の大部分は27-20億年前に形成されたものですが、7億年前にも大規模な縞状鉄鉱床が形成されました。縞状鉄鉱床ができるためには無酸素の海中に鉄イオンが溶け込むことが必要です。すでに酸素の十分にあった時期にどうして縞状鉄鉱床が生成されたかについては、大氷河期(スノーボール・アース)との関連が指摘されています。
全世界の鉄鉱石埋蔵量1500億トンのうちロシア、オーストラリア、ウクライナ、中国、ブラジルの上位5カ国だけで約3/4を占めています。鉄資源について考える場合、鉄鉱石の埋蔵量=鉄埋蔵量とはならないことに注意しなければなりません。鉄鉱石埋蔵量 X 品位(鉄含有率)が実際の鉄の埋蔵量ということになります。
世界の鉄鉱石埋蔵量は1500億トンですが現在は商業ベースに乗らないものを含めると1兆トン程度と見積もられています。市場経済においては資源は良質なものから採取されるという原則が成立します。
中国は少し特殊な事情もありますが、世界の製鉄会社はまずブラジル、オーストラリアなどの高品位の鉄鉱石を利用したいと考えます。商業的に価値のある鉄鉱床としての条件は次のようになります。
(1)地表から浅いところにあり採掘しやすいこと
(2)鉱床の規模が大きいこと
(3)品位(鉄含有率)が高いこと
実際には粗鋼を生産する国への運搬のしやすさも重要な要素となります。鉄鉱石を採掘し運搬するためにはコストがかかります。品位が半分の鉄鉱石を利用すると、同じ粗鋼を生産するのに2倍の鉄鉱石が必要となります。これは、粗鋼の生産コストに直接響いてきます。
中国は世界最大の鉄鉱石生産国ですが、同時に世界最大の輸入国でもあります。現在の中国は世界の1/3の鉄鋼を消費しています。世界の鉄鉱石の埋蔵量は年間生産量(採掘量)に比して70倍程度ですが、高品位の鉄鉱石は枯渇が心配される時期にさしかかっています。
それでも、金属資源の中で鉄資源はゆとりのある部類に入ります。中国の輸出規制で脚光を浴びるようになったレアメタルを例にとるまでもなく、ベースメタルの可採年数(可採埋蔵量/年間採掘量)は銅(31年)、スズ(22年)、鉛(20年)、金(17年)、銀(14年)とものによってはレアメタルより少ないものもあります。
社会生活に欠かせない資源でもっとも深刻なものは銅資源でしょう。中国、インド、ブラジルなど人口大国が経済成長路線を進むと銅消費量は急増し、可採年数は急速に小さくなることでしょう。
右の画像上はハマースレイ縞状鉄鉱床、画像は「岐阜大学教育学部・地学教室」より引用しました。西オーストラリアの北西部に分布するハマースレイ鉄鉱床は24億年前に広大な大陸棚に堆積しました。
右の画像下は縞状鉄鉱床地層を拡大したもの、画像はwikipedia より引用しました。酸化鉄の層と泥の層が交互に積みあがっています。
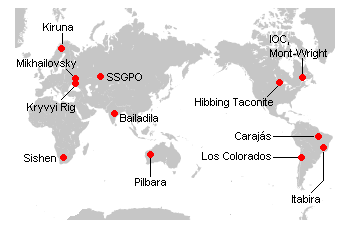
世界の主要鉄鉱山、画像は「
DGX100・資源源について」から引用しました。ここに示される鉱山はすべて27-20億年前に形成された縞状鉄鉱床に由来します。
| 国名 | 鉄鉱石 埋蔵量 |
鉄 埋蔵量 |
鉄鉱石 生産量 |
|---|---|---|---|
| 中国 | 21,000 | 7,000 | 770 |
| ブラジル | 16,000 | 8,900 | 390 |
| オーストラリア | 16,000 | 10,000 | 330 |
| インド | 6,600 | 4,200 | 200 |
| ロシア | 25,000 | 14,000 | 110 |
| ウクライナ | 30,000 | 9,000 | 90 |
| 米国 | 6,900 | 2,100 | 54 |
| 世界全体 | 150,000 | 73,000 | 2,200 |
世界の鉄資源と鉄鉱石生産量(2008年)、単位は100万トン、出典はUSGS(アメリカ地質調査所)。鉄埋蔵量は鉄鉱石埋蔵量に品位(鉄含有率)を乗じた数値と思われます。世界の優良鉄鉱床の平均品位は65%にもなりますが、中国産鉄鉱石の品位はその半分程度と推定されます。
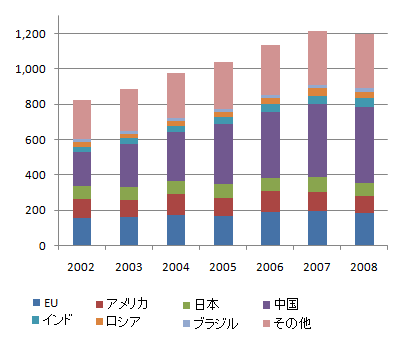
世界の粗鋼消費量の推移、単位は100万トン、出典はUSGSと思われます、画像は「投資日記」から引用しました。中国が世界の1/3を占めており、21世紀に入ってからの世界の消費量の増加分はほとんど中国によるものです。
多細胞生物の出現(10億年前)
多細胞生物とは動物、植物、菌類に分かれており、それぞれ独立して多細胞化したと考えられます。真核生物が誕生したのはおよそ20億年前、多細胞生物が誕生したのは10億年ほど前と考えられています。最近はこの起源が15億年前にさかのぼるのではという報告もあります。
原生生物(単細胞の真核生物)は細胞分裂がそのまま固体の増加につながりますので、種の維持・繁栄という観点からは有利と考えられます。多細胞生物の大半は有性生殖で繁殖しますので、繁殖速度は原生生物に劣ります。
しかし、多細胞生物は細胞を専門化させ、複雑な機能を獲得することにより生存を有利にする戦略をとってきたと考えられます。実際に地球上では数はともかく、多細胞生物が進化の頂点に君臨しています。
原生生物から多細胞生物である動物、植物、菌類の進化はそれぞれが独立していたと考えられています。そして、少なくとも植物に関しては複数回の多細胞化への試行があったことも確認されています。
単細胞生物が多細胞生物に進化するためには、細胞同士の接着や周囲の細胞との協調が必要です。このような機能を獲得するためおよそ10億年という時間がかかったと考えられています。
最初の動物(多細胞動物)はカイメンであると考えられています。カイメンは複数種類の細胞から構成されており、その中に襟細胞があります。襟細胞は一層にならんで袋状の襟細胞室を作ります。
襟細胞室はカイメンの体中に張り巡らされた水管(水の通り道)の壁とつながっています。襟細胞室の中には1本の鞭毛(べんもう)が付属しており、この鞭毛が動くことで水流を起こし、その流れに乗ってくる栄養粒子を襟細胞が取り込みます。襟細胞は他の細胞に栄養素を渡しているので原始的な消化器官の役割を担っています。
この襟細胞の機能は原生生物の襟鞭毛虫の食べ物の摂り方と非常によく似ています。襟鞭毛虫の体長は最大でも10μm程度であり、一本の鞭毛とその基部に微絨毛が環状に取り囲んだ襟状の構造をもっています。鞭毛は水流を起こして栄養粒子などを集め、これを襟によって捕捉して食べます。葉緑体はもっておらず、完全な従属栄養型の生物です。
襟鞭毛虫は生活環の一時期に300-500μmほどもある大きなコロニー(群体)を作り、しかもコロニーの中心部の個体は形態が変化することも知られています。外縁部の個体は本来の姿を維持していますが、中心部の固体は球形となり襟や鞭毛が発達しません。襟鞭毛虫のコロニーの中では個体(細胞)の分化に近いことが起きています。また、変形動物やワムシなどにも襟鞭毛虫に似た細胞がみられます。
単細胞生物から多細胞生物の進化過程において重要な要素は細胞の接着です。そのためには、細胞同士は密着するが融合や食作用は起きないような特殊な接着分子(カドヘリン)が必要になります。このカドヘリンの元となる基質が襟鞭毛虫にも見られます。
このようなことから襟鞭毛虫が多細胞動物の起源であるという学説が広く支持されています。襟鞭毛虫のコロニーが個体の変化を生み出すことから、多細胞動物は襟鞭毛虫のコロニーが固定化され、全体でひとまとまりの生物に進化していったというストーリーを描くことができます。真核生物の進化は真正細菌との共生によりもたらされたように、多細胞動物への進化は同種の細胞の共生によりもたらされたということができます。
そして、起源となる襟鞭毛虫にもっとも近いカイメンが多細胞動物の始まりであると考えられています。カイメンに始まる多細胞動物を総称して後生動物(metazoans)と呼びます。これが襟鞭毛虫起源説の概要です。
現在の系統分類ではカイメンは器官が明確に分化していないことからカイメンを除く後生動物をまとめて真正後生動物としています。また、背骨をもつ動物(脊椎動物)と背骨をもたない動物(無脊椎動物)に分ける2分法もありますが、こちらはヒトに近い種を基準にする便宜的なものです。そもそも脊椎動物という門はなく脊索動物門に含まれています。
多細胞生物の出現は生物の複雑化、大型化への道を開き、およそ9億年前に始まったとされる有性生殖という新しいシステムを採用することにより、やがて多彩な生物へと進化していくことになります。
しかし、地球の極端な寒冷化により生物の危機が訪れます。現在の太陽光度でも二酸化炭素がゼロになると地球の平均気温-18℃にもなります。10億年前の太陽光度は現在よりも10%低かったので、代表的な温室効果ガスであるメタン、二酸化炭素の減少は大氷河時代を引き起こすことになります。
地質時代を通して大気中のメタンや二酸化炭素の量は大きく変動しており、それにより地球表層気温も高温側にも低温側にも大きく振られています。
海水中の二酸化炭素は陸地の風化により運ばれる陽イオンとの結合により、一定の速度で消費されます。また、化石燃料のように生物に取り込まれ固定化されて消費されます。これを補うように火山などから二酸化炭素が供給されます。ところが、地球内部から供給される二酸化炭素が減少するとすると温室効果が少なくなり、地球は寒冷化します。
地球が極端に寒冷化する大氷河時代は地球史おいて複数回、少なくとも2回、ヒューロニアン氷河時代(約24億5000万年前から約22億年前)の最終期と、マリノニアン氷河時代(約7億3000万年前~約6億3500万年前)に発生しています。全球凍結あるいはスノーボールアース仮説といい、現在では多くの証拠が見つかり、広く受け入れられています。
23億年前の大氷河時代はその頃の温室効果ガスの主役であったメタンガスが、大気中に増加した酸素と結合して減少したことによるとされており、7億年前には二酸化炭素の減少がその要因と考えられています。
現在は300万年前に始まった氷河期にあたり、北半球の氷床が成長する氷期と後退する間氷期が交互に発生しています。最後の氷期は1万年前に終わり、現在は間氷期にあたります。しかし、大氷河期はこの300万年間の氷河期とはスケールが違います。
熱帯地域を含め地球の表面はすべて氷床で覆われていたのです。海氷の厚さは1000mにもなりました。地表が氷雪に覆われるとアルベド(地表の反射率)が高くなり、太陽光を反射するため、地表は急速に寒冷化していきます。そして、ひとたび地球の全表面が氷床で覆われると、再び液体の海には戻らないと考えられてきました。
実際、全球凍結は数千万年から1億年以上も続いたと考えられています。しかし、地球内部と表層間の物質循環により二酸化炭素が供給され続け、大気中の二酸化炭素が現在の500倍くらいになると気温が上昇し、一気に氷床は融けたとされています。
大氷河時代の間、生物が生き延びられる場所は火山の周辺などに限定され、原生生物の大量絶滅が発生しました。全球凍結が終了すると地球は温暖化し、生物は新しい環境で新たな進化を遂げることになります。
22億年前には真核生物が繁栄し、6.3億年前にはシアノバクテリアに代表される藍藻類の大発生により海水中の酸素濃度は急激に増加します。真核生物の一部はこの豊富な酸素を利用し、細胞接着(構造)物質であるコラーゲンを産生することに成功し、エディアカラ、カンブリアにつながる多様な大型多細胞生物が出現するようになりました。
地球の全球凍結、あるいは複数回発生した生物の大量絶滅の度に生物は新しい環境に適応した進化をとげ、大量絶滅以前にも増して豊かで多様な生態系を形成してきました。逆に、大量絶滅のような極端な試練がなければ、生物の進化はまったく別なものとなったと考えられます。
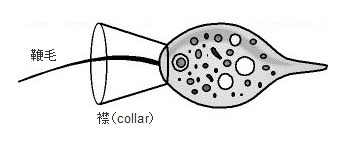
襟鞭毛虫(えりべんもうちゅう、Choanoflagellida)、画像は「原生生物図鑑・襟鞭毛虫目」より引用しました。原生生物の中でもっとも後生動物(現在の多細胞動物)に近いと考えられています。体長は最大でも10μm程度であり、一本の鞭毛とその基部に微絨毛が環状に取り囲んだ襟状の構造をもっています。鞭毛は水流を起こしてバクテリアなどを集め、これを襟によって捕捉して食べます。葉緑体はもっておらず、完全な従属栄養型の生物です。


ヘッケルの描いたカイメン(上)と自然の芸術図形(下)、画像はwikipedia から引用しました。エルンスト・ヘッケル(1834-1919年)はドイツの生物学者であり、哲学者です。ヘッケルは「個体発生は系統発生を反復する」という「反復説」を提唱し、その研究から多細胞動物は鞭毛虫の群体から生まれたとするガスツレア説を主張しました。彼の生物スケッチはまったく細密で、ほとんど芸術作品のようです。カイメンのスケッチがあまりにも素晴らしかったので、「自然の芸術図形」からもう1枚引用することにしました。

全球凍結した地球の想像図、画像はwikipediaより引用しました。現在、判明している範囲ではこのような全球凍結は29億年前、24億年前、7億年前に発生したと考えられています。
カンブリア爆発(5.5億年前)
古生代の始まりにあたるカンブリア紀(5.42-5.1億年前)は多細胞動物が大きく発展した時期でカンブリア爆発とも表現されています。しかし、多細胞生物の発展はカンブリア紀の前のエディアカラ紀(6.4-5.42億年)から始まっていたようです。
もっとも、少し前の地質時代区分では化石が基準となっていましたので、標準化石がない原生代などは約20億年がひとくくりとなっています。さすがにそれではまずいというので、現在は原生代が新原生代(5.42-9.00億年前)、中原生代(9-16億年前)、古原生代(16-25億年前)に区分され、さらにそれぞれが細分化(紀に相当します)されています。
「wikipedia の地質時代」のページには目で追うのも大変なくらい細かい区分が載せられています。ここの注意書きには「過去四半世紀の研究で先カンブリア時代の地質学的・古生物学的な見方が大きく変化したが、そもそも地質学的証拠に乏しい時代であるため、年代区分を定義する正確な用語はまだ定まっていない」と記載されています。
エディカラ紀は新原生代最後の時期に相当します。この時期の化石がたくさん見つかるようになったので、原生代と古生代の境界時期も変更されたようです。古生物学的な新しい発見と地球の歴史の新しい知見により、原生代以前の地質時代区分も更新されています。
この時期をエディアカラというのはオーストラリアのエディアカラ丘陵で多数の化石が発見されたことによります。同時期の化石はカナダのニューファンドランド島、ロシアの白海沿岸、南アフリカ、中国南部などでも発見されています。これらの化石採集地を代表してエディアカラが地質時代の名称となっています。
この時期に発見された化石はエディアカラ動物群と呼ばれており、固い殻(外骨格)をもたないことと非常に扁平であるという特徴をもっています。大きさは数十cmもあるにもかかわらず厚さは数mmほどしかありません。
この特異な形状から一時期、エディアカラ動物群はカンブリア紀に登場する多様な動物には結びつかない、傍系のものとも考えられましたが、現在ではカンブリア動物群の祖先であると考えられています。それでも、この時期に動物の大分類のあたる門に相当するような新しい生物が生まれ、絶滅したことは確かなことにようです。大氷期が終了して多細胞生物の大発展の序章に相当するのがエディアカラ動物群ということです。
エディアカラ動物群の化石は肉眼で観察できる最古のものです。硬い殻(外骨格)をもたない生物が化石として保存されることは非常に珍しいことです。にもかかわらず世界中でそのような化石が発見されるということは、この時期に肉食あるいは腐食性(死骸を分解する)生物が少なかったと考えられます。
柔らかい生物の化石が保存される例外的な事例は大量の土砂により生物が一気に低酸素の状態で埋まってしまった場合です。しかし、そのような場合に形成されるのは頁岩(泥岩)となります。
エディアカラ紀に続くカンブリア紀の化石がよく産出されることで有名なカナダのバージェス頁岩はそのような仕組みで多くの生物を泥の中に閉じ込めてしまいました。そのため、バージェス頁岩からはまるでタイムカプセルのように保存状態の良い化石がたくさん見つかります。
エディアカラ動物群の化石は砂岩や粗粒泥岩などの堆積物から見つかることが多いのです。このような堆積層は流れがあり、酸素の豊富な海底で形成されるため、柔らかい組織をもった動物の保存には適していません。このような観点からエディアカラの時期には捕食動物が存在せず平和な楽園だったのではと考えられます。
エディアカラ動物群は海底に固定されていたり、海底をゆっくり這いまわったり、水中を浮いたりしながら、水流で運ばれてくる栄養粒子を取り込んでいたようです。肉食性の動物がいなかったので、固い殻をもたなくても問題はなかったわけです。
残念ながらエディアカラ動物群の大半は古生代の始まり以前に絶滅しています。5億5000万年前に超大陸ゴンドワナが形成されましたので、それに起因する火山活動がエディアカラ動物群の絶滅の原因ではと考えられています。また、カンブリア紀に登場した肉食性の動物により、あっさり駆逐されてしまったという考えも否定できません。ともあれ、動物進化の舞台はその後のカンブリア紀に引き継がれたわけです。
カンブリア紀(5.42-5.10億年前)はおよそ3000万年の短い期間ですが後生動物(現在の動物のことです)にとっては最大のイベントとなった時期でした。温暖で浅い海の広がる環境、十分な酸素のおかげで動物進化史上最も重要な出来事が起こりました。
それまで数十数種しかなかった動物が短期間のうちに数万種にまで爆発的に増加しました。そのためこの進化の大イベントは「カンブリア爆発」と呼ばれています。現在の後生動物は36の門に大分類されていますが、カンブリア紀にはその多くの門の祖先が出そろっています。中には現在の門やその下位分類に含めることのできないものも多数存在しています。
カンブリア紀の化石の主要産地は2か所あります。一つはカナディアン・ロッキーの山中にあるバージェス頁岩(5.05億年前)、もう一つは中国雲南省澄江県(5.25億年前)の地層から出土するものです。それぞれの化石の生産地にちなんでバージェス動物群、澄江動物群と呼ばれています。
カンブリア紀の多彩な動物群が明らかになったのはバージェス頁岩というすばらしい化石の宝庫が露頭していたおかげです。頁岩とは目の細かい粘土状のシルトが堆積した泥岩の一種で、層状になっているためまるで本のページをめくるように薄く剥離することから頁岩と呼ばれています。バージェス頁岩の化石にには固い殻をもった動物のみならず、体の柔らかい部分まではっきりと残されています。
このことから、バージェス頁岩は斜面の下にある200mほどの深さの海底で堆積したものと考えられます。このような地形のところではときどき斜面から土砂が崩落し、乱泥流により海底の生物が一気に泥の中に閉じ込められことが起こります。酸素の少ない泥の中に閉じ込められた生物は柔らかい部分や細かい部分もきれいに残されたわけです。
バージェス頁岩を発見したのは米国の古生物学者であるウォルコットです。彼は1910年から数次にわたる発掘を行い65,000個を超える化石を収集しました。バージェス頁岩のあるカナディアンロッキーの山中は厳しい寒さと降雪のため、夏のほんの一時期しか発掘はできませんので、発掘は数シーズンにわたることになります。
彼は化石となった動物は現存する分類群の祖先的形質を多くあらわしたものと考えていたようです。この時期はカンブリア紀のもっとも良質な化石が発見されたこと以上の脚光を浴びることはありませんでした。
しかし、1960年代後半よりウィッチントン(英国人、ハーバード大学古生物学教授)を中心とする研究グループがこれらの化石の再調査を行ない、発見された化石動物が既存の分類群に当てはまらないものが多く、中には門そのものの帰属すらはっきり確定できないものが多数あることを明らかにしまた。これにより、それまでのカンブリア紀動物に対する古生物界の認識が一変しました。
カンブリア動物群を一般の人々に知らしめたのはグールド(米国の古生物学者、進化生物学者、ハーバード大学比較動物学教授)が1989年に出版した「ワンダフル・ライフ(バージェス頁岩と生物進化の物語)」です。
このベストセラーとなった著書の中でグールドはカンブリア動物の驚くべき多様性とその進化に対する考え方を記しています。グールドは「カンブリア紀動物群の多様性が最大点であり、その後はそれを超えることがなかった。従来のダーウィン的な進化(小進化)とは異なるメカニズム(大進化)がある」と主張しました。この仮説は断続平衡説と呼ばれており、広く議論を引き起こしました。
実はグールドの主張はウィッチントンのもとで研究を進めたコンウェイ・モリスの初期の考え方に影響されたものでしたが、その後、モリスはその著書「カンブリア紀の怪物たち」でグールドの考え方を批判しています。モリスは一見非常に奇妙に見える動物でも既存の門の分類に含めることができると記しています。
もっとも、カンブリア紀に多数存在していた節足動物の中には現在につながらない種が多数存在していたことはモリスも認めています。カンブリア紀動物群は当時の環境で行われた進化の大実験でした。
すばやく動くことのできる捕食動物の出現が淘汰圧力を強め、進化を促進したことも多様性の一因だったと考えられています。弱い動物は捕食動物から逃れるため、いろいろな手段を講じました。多くの生物のデザインが試され、その中で周囲の環境(捕食・被捕食の関係も含みます)に適応したものが次の系統の祖先になりました。

地質時代区分の一例、単位は1億年。時代区分の数字は出典により微妙に異なります。多細胞動物が大発展したカンブリア紀の前にエディアカラ紀があります。エディアカラ紀はその前の大氷期が終了して多細胞生物が発展期に入った時期です。
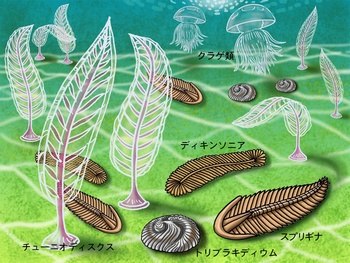
エディアカラ生物群、画像は「DINO-KEN・恐竜博士に聞こう」から引用しました。エディアカラ生物の大半はカンブリア紀までには絶滅しましたが、一部はカンブリア紀の系統につながっていると考えられます。
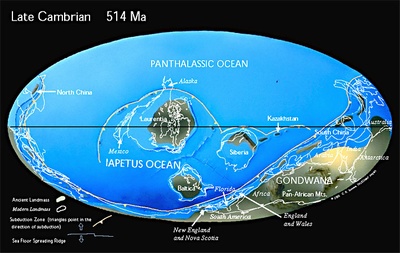
カンブリア紀の大陸配置、画像は
「PALEOMAP Project」より引用しました。超大陸ロディニアが分裂し、5.5億年前に超大陸ゴンドワナが形成されました。この時の火山活動によりエディカラ動物群の多くは絶滅し、仕切り直しの形でカンブリア紀の進化の爆発が始まります。

バージェス頁岩の採掘地(ウォルコット採石場)、画像はwikipedia より引用しました。バージェス頁岩の露頭はカナダのブリティッシュコロンビア州にあります。バンフの近くにある観光地のエメラルド湖(レイククルーズ村が湖に面しています)からバージェス峠を経由し、フィールド山から奥のワプタ山に向かうところに位置しています。

カンブリア紀の動物を記した「ワンダフル・ライフ(グールド著)」と「カンブリア紀の怪物たち(モリス著)」は日本語でも出版されています。ワンダフル・ライフの方がセンセーショナルな語り口ですので一般受けするでしょう。
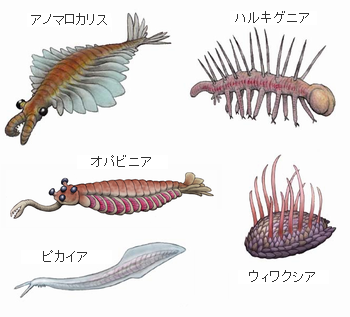
バージェス動物群、画像は「古世界の住人」より引用し編集しました。確かに現在の動物門のどこに入れたらよいか迷うようなものがたくさん存在していたようです。アノマロカリスは最大で2mにもなり、この世界の最強の捕食者でしたが絶滅してしまいます。アノマロカリス以外のものは10cm以下の大きさです。固い外骨格をもたないピカイアは生きのび、軸索動物(脊椎動物)の祖先になりました。
オゾン層の形成(5億年前)
この時期に大気中の酸素濃度も現在に近い21%となり、オゾン層が形成されます。オゾン層は成層圏に存在し、高度20-25kmくらいのところがもっとも高密度となります。大気中に酸素が増加すると成層圏では紫外線により酸素分子が分解され酸素原子が作られます。この酸素原子は酸素分子と結合するとオゾン(O3)となります。
オゾンは非常に酸化力が強く、殺菌・消毒に用いられることからもわかるように生物にとっては有害物質です。光化学スモッグ注意報が発令されるときの指標の中にはオゾンも含まれています。
地表ではやっかいもののオゾンですが成層圏にあるオゾン層は生物にとって重要なバリアとなっています。オゾン層におけるオゾン濃度は2-8ppmであり、垂直方向にすべてのオゾンを集めても0℃、1気圧では3mmほどにしかなりません。しかし、このオゾン層は波長の短い紫外線を吸収するという重要な働きをもっています。
人間の眼が光として感じることができる波長は400-770nmです。これより波長の短いものを紫外線(UV : ultraviolet rays)、波長の長いものを赤外線(IR : infrared rays)といいます。
太陽光に含まれる紫外線は波長によりUV-a(長波長紫外線、波長320-400nm)、UV-b(中波長紫外線、波長290-320nm)、UV-c( 短波長紫外線、波長 200-290nm) の三種類に分けられます。波長の短い紫外線ほどエネルギーが大きいので生物にとっては有害です。
オゾン層はUV-c(短波長紫外線)を100%、UV-b(中波長紫外線)を90%ほど吸収しますので、生物にとっては重要なバリアとなっています。それでも波長の長い紫外線にしばらくあたるだけで日焼けをします。海辺のような紫外線の強いところでは強い日焼けにより皮膚が損傷を受けます。また、雪原も紫外線が強いためサングラスなどで眼を守る必要があります。
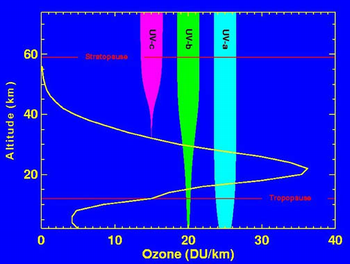
オゾン層の働き、画像はwikipeda より引用しました、縦軸は高度、横軸はオゾンの密度を表しています。大気中の酸素が紫外線により活性化され、高度20-25kmあたりのところにオゾン層が形成されます。オゾン層では波長の短い紫外線が吸収され、生物にとって有害な紫外線が地表に届かなくなります。オゾン層というバリアができたので生物は陸上に進出できるようになりました。
生物の大繁栄(5.1-2.45億年前)
カンブリア爆発を経て、生物の進化は加速します。細胞内にある核という器官に大量の遺伝子を貯蔵できるようになったことがその要因としてあげられます。
細菌類は生存に最小限必要な遺伝子しか持たなかったため、劇的な変化をすることなく現代に至っています。それに対して、真核生物は現在必要としない遺伝子をどんどん取り込むようになり、進化の可能性の範囲を広げました。
オルドビス紀(5.1-4.39億年前)を代表する動物はオウムガイ(軟体動物)、三葉虫(節足動物)、筆石(半索動物)、甲冑魚(魚類)などです。オウムガイの仲間は現存してしており、生きた化石の一つです。甲冑魚は現在の魚類の祖先にあたります。
オゾン層ができたことにより、生物は安心して海から上陸できるようになりました。植物はシルル紀(4.39-4.08億年前)に上陸を果たしたとされてきましたが、これは化石の発見により5億年前にさかのぼる可能性があります。節足動物の上陸もシルル紀とされています。
脊椎動物の上陸はデボン紀(4.08-3.63億年前)とされています。脊椎動物が陸に上がるためにはいくつかの準備が必要です。最大の問題はミネラルの補給と重力でした。
海には生物に必要なミネラルが豊富に存在していますが、陸上ではそうはいきません。脊椎動物は骨にミネラルを貯蔵することによりこの問題をクリアしました。骨格は動物の体を支える働き以前にミネラルの貯蔵庫だったわけです。
骨に含まれるリン酸カルシウムは溶出と生成が繰り返され、体内の濃度を一定に保っています。体内にカルシウムが不足すると骨からカルシウムが供給されます。そして、カルシウムが余ると骨が生成されます。
最初に上陸を果たした脊椎動物はユーステノプテロンという肉鰭綱の魚であったろうと考えられています。ユーステノプテロンは体長は30-120cm、潟湖などの酸素の少ない環境に適応するため原始的な肺呼吸のシステムをもっていました。
それは、食道の一部を膨らませ、現在の肺魚が行っているように水面に口を出して空気を飲み込み、肺から酸素を取り込んでいました。その後、肺呼吸の不要な川や海の環境に適応した魚は、肺を浮き袋に進化させています。また、肉鰭綱の魚は植物の多い浅い海底を移動するため4つの胸鰭と腹鰭の内部に硬い骨を発達させました。
最初の陸上脊椎動物といわれているイクチオステガの化石は3.6億年前のものでした。イクチオステガは頑丈な4本足と肋骨を備えており、水辺の環境で暮らしていたと考えられています。このイクチオステガがすべての陸上脊椎動物の祖先になります。
石炭紀(3.63-2.9億年前)になると巨大なシダ植物が水辺に高さ20-30mにもなる大森林を作りました。幹の太さは最大2mにもなります。現在のシダ類の大半は林床のように日のあまり当たらないところで小さな姿で生き残っています。それは他の裸子植物や2億年前頃に誕生した被子植物との競合に負けた結果でしょう。
それでも、木性のシダは現在でも熱帯や亜熱帯地域に生き残っており、高さは5-10mくらいにもなります。多くの木性シダは茎の上部に成長点をもちヤシのように大きな葉を茂らせます。
このような葉は幹から直接伸びる葉柄により支えられています。葉が老化すると葉柄の根元から脱落して幹にその葉痕が独特の鱗状紋となります。小笠原の固有種であるマルハチ(ヘゴ科・ヘゴ属)は葉痕が丸に囲まれた上下が逆になった漢字の八の字に見えることからマルハチと命名されました。
石炭紀の木性シダもこのような形態であったと考えられます。代表的なものはリンボク、フウインボク、カラミテス、プサロニウスです。フウインボクの幹にある葉痕は六角形の形をしていますし、リンボクの幹はうろこのようになっていることからこの名前が付けられました。
このようなシダ植物の森林には巨大な節足動物が生息していました。全長60cmもある巨大なウミサソリ(メガラシネ)、翼長70cmの巨大トンボ(メガネウラ)、全長2mの巨大ムカデ(アースロプレウラ)などの化石が残されています。
1年中湿潤な熱帯気候と豊富な酸素(陸上植物の繁栄により31-35%ほどもあったと考えられています)が節足動物の巨大化を後押ししたようです。肺をもたない種類の節足動物は気管を通して拡散により酸素を取り込んでいます。大気中の酸素濃度が高いと拡散が容易となり、同じ表面積の気管でたくさんの酸素が利用できるからです。
逆に大気中の二酸化炭素濃度は現在(約350ppm)の4倍程度に減少しています。これは、大きな火山活動がなく陸上の植物に大量の二酸化炭素が固定されたためです。したがって、石炭紀は寒冷化に向かう時期ということになります。
石炭紀はその名の通り石炭が大量に生成された時期です。水辺の湿地に大森林が形成され、それらが埋没して石炭のもととなったとされています。しかし、なぜこの時代の森林だけが巨大な石炭層を形成できたのか興味のあるところです。
この時代の森林は胞子で繁殖するため水辺だけに限定されていたはずです。現在のように(環境が許せば)大陸全体に森林が広がっている時代とはだいぶ状況が異なっているはずです。樹木の蓄積量を比較すると現在の方がずっと多かったのではと推測します。
にもかかわらず石炭層を形成できたということは、死んだ樹木が非常に効率的に泥炭化したと考えられます。現在の熱帯の森林は石炭ができるような環境にはありません。倒れた樹木は速やかに分解されて土に還るからです。一方、比較的低温の湿地帯では分解速度が遅いので泥炭が効率よく形成されます。
こう考えると石炭紀の環境は樹木を分解する生物に乏しかったか、そのような生物の分解能力が低かったと考えられます。実際、現在の菌類や真菌類が担当しているように、樹木の主要構成物質であるセルロースやリグニンを分解するのはとても大変です。石炭紀に大繁栄した木性シダは新しい生物のため、まだ分解系の生物が十分に進化していなかったのではと考えられます。
世界の石炭の全埋蔵量(可採埋蔵量ではありません)は3.4兆トンと見積もられています。石炭に占める炭素の重量割合は70-80%程度です。平均して75%として計算すると、世界中の石炭に含まれる炭素は2.6兆トンとなります。
現在の大気中の(二酸化炭素として存在する)炭素は7500億トンですらか、石炭の中に固定されている炭素はその3.5倍ということになります。二酸化炭素は大気と海の間でやりとりされており、大気中の二酸化炭素分圧が下がると海から供給されますが、これほどの炭素が固定されると地球表層の二酸化炭素は減少することになります。
石炭紀が始まる前は現在の8倍近くあった大気中の二酸化炭素は石炭紀の間に4倍程度まで減少しています。このため、石炭紀の終わりからペルム紀末までは寒冷な気候となりました。石炭紀の平均温度は22℃、その後の寒冷期の平均気温は13℃程度です。ちなみに、現在は氷河期の間氷期にあたり平均気温は15℃、氷期の最盛期は9℃ほどになります。
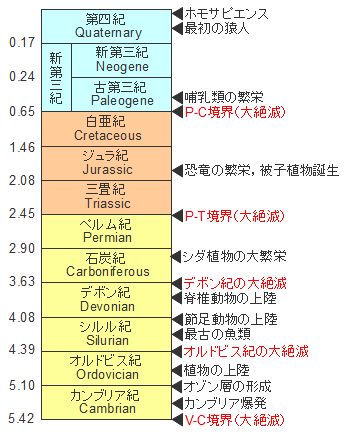
生物の大繁栄、縦軸の単位は億年です。カンブリア爆発を経て生物は海中と陸上で大繁栄します。しかし、地球は絶え間なく変化しており、環境の激変は生物の大絶滅を引き起こします。
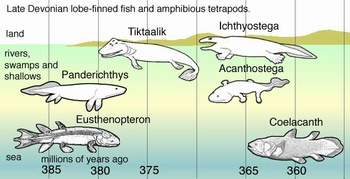
デボン紀の肉鰭類と四足動物、画像はwikioedia から引用しました。ユーステノプテロンとイクチオステガの間に中間的な動物も描かれています。図中の「Coelacanth」は生きた化石といわれているシーラカンスの仲間です。

シーラカンス(Latimeria chalumnae)、画像はwikipedia より引用しました。シーラカンスの仲間は3.8億年前に誕生し、白亜紀末には絶滅しています。しかし、2種だけは当時とほとんど変わらない姿で現在も生き残っており、生きた化石と呼ばれています。現生種はシーラカンス目・ラティメリア科・ラティメリア属に含まれています。このため現生種の正式名称はラティメリアとなります。
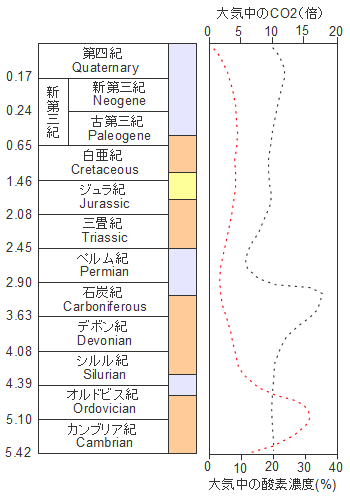
古生代。中生代・新生代の気候と大気中の酸素と二酸化炭素濃度、薄いオレンジの時期は温暖期(平均気温22℃)、青みがかった灰色は寒冷な時期(平均気温13℃)を表しています。赤の点線は大気中の二酸化炭素濃度で現在(約370ppm)に対する倍率で表示しています。暗い灰色の線は大気中の酸素濃度を表しています。
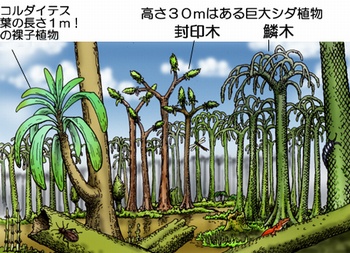
石炭紀の世界、画像は「古代世界の住人」「古代世界の住人」から引用しました。水辺の湿地には高さ30mにもなるシダ植物が繁茂し、巨大な節足動物が暮らしていました。

石炭紀の巨大昆虫メガネウラ、画像は上と同じ「古代世界の住人」から引用しました。ここのサイトにはお仕事で描かれた古代世界の生物の素晴らしい画像が数多く掲載されています。このページでは3点の画像を引用しています。業務で制作した画像ですので著作権に抵触するかもしれません。ご指摘があれば削除します。メガネウラは地球史上最大のトンボで翼開長は60-70cmくらいあったとされています。
恐竜の大繁栄(2億年前)
古生代はペルム紀で終わり中生代(2.45-0.65億年前)が始まります。この境界(P-T境界)で地球史上最大規模とも言われる大量絶滅が起こりました(ベルム紀の大絶滅)。このとき、海洋生物のうちの96%、全ての生物種の90%から95%が絶滅したとされています。
この原因と考えられているのがシベリア洪水玄武岩(シベリア・トラップ)の噴出であり、それを引き起こしたのは超大陸パンゲアを分裂させたスーパーホット・プルームであると考えられています。ただし、隕石の衝突や他の原因とする説もあり、議論は決着していません。
大陸分裂の原動力となるスーパーホット・プルーム(下部マントルと上部マントル間の対流)は下部マントルと上部マントル境界付近ででいったん滞留します。ところが、ときにはそのまま上昇して地殻を破って噴出することがあります。これは地球上でもっとも激しい火山活動になります。
スーパーホット・プルームの巨大な高温物質は2.5億年前にロシア東北部・中央シベリア高原を中心に噴出し、その結果、現在わかっている範囲でも200万km2の範囲に洪水玄武岩層が形成されました。隠されている部分を含めると400万km2にもなるとされています。噴火は100万年にわたり何回か発生し、その総量は富士山が過去1万年間で噴出した溶岩量の10万倍程度と見積もられています。
火山灰を吹き上げる爆発的な噴火ではありませんが、この火山活動により大量の火山ガスが放出され、地球の気候は激変します。また、海底のメタンハイドレードの相当部分が崩壊して大気中に大量のメタンが放出され、それが酸素と結びつくことにより、大気中の酸素は半減しています。この激しい火山活動とその後の気候の激変が生物種の大絶滅を招いたと考えられています。
中生代の最初にある三畳紀の地球環境は平均気温23℃、大気中の酸素濃度が12-15%というものでした。大絶滅によりペルム紀で繁栄していた哺乳類の祖先である単弓類の大部分は絶滅しました。海では三葉虫が絶滅しています。ちなみに、ヒトは酸素濃度が18%未満になると酸欠状態になります。15%以下(ろうそくが消える)では息苦しく、力が入らなくなる危険な状態になります。
三畳紀の陸上生物の主役を決めたものは乏しい酸素であったとされています。大気中の酸素濃度の低い状態は約1億年続きました。この乏しい酸素環境で繁栄を開始したのは恐竜の仲間でした。比較的小型だった恐竜は種を増やし、酸素が増加した三畳紀末期から急速に大型化していきます。
恐竜の繁栄は気嚢システムをもったことと関係しているようです(恐竜気嚢仮説)。気嚢は現在の鳥類がもっているすぐれた呼吸システムであり、それは恐竜から受け継いだものと考えられます。気嚢システムにより鳥類は飛翔という高い運動能力をもち、酸素の少ない数千mの高度を飛ぶことができるのです。
鳥類の呼吸器は大ざっぱに後気嚢、肺、前気嚢に分かれています。外気はまず後気嚢のポンプ作用で吸気され、肺に送られ、前気嚢のポンプ作用により肺から出され排気されます。肺の中の空気の流れが一方向であるため、肺にはいつも新鮮な空気が供給されされます。この仕組みは肺の中で吸気と呼気が混ざり合う哺乳類のものに比べてずっと効率が良いのです。
恐竜(一部もしくはすべての)はこの気嚢システムをもつことにより、酸素の乏しい環境でも繁栄し大型化できたと考えられています。恐竜以外の動物も呼吸システムを改良しています。哺乳類は肺と横隔膜により、ワニは横隔筋により呼吸量を増やしています。
呼吸量だけではなく血液の循環量も増やす必要があり、ワニ・恐竜・鳥類・ほ乳類の心臓は2心房2心室に進化しています。環境の悪化は進化の原動力となっているわけです。恐竜は中生代を通して1.5億年にわたり繁栄しましたが、それはこの時代の環境が比較的安定していたことが大きな要因です。
生物の系統分類による定義では恐竜とは陸上を歩くものに限定されています。イルカにそっくりな魚竜、4本のひれのような足で水中に暮らしていた首長竜、空をグライダーのように滑空した翼手竜などは恐竜の中に入りません。これはちょっと残念な分類ですね。
もう一つ、系統分類においては鳥類の扱いをどうするかが問題になります。鳥類の祖先は爬虫類か恐竜かは長い間の論争になっていましたが、どうやら恐竜ということで決着がつきました。現生生物の分類では鳥類は爬虫類から独立していますが、系統分類で追うと恐竜の末端に位置するようになります。これも、生物学者の悩みの種になっています。
そもそも爬虫類というグループでまとめてしまうのは無理があるのではという議論もでています。化石を含め爬虫類は単弓類(哺乳類はここに含まれます)と双弓類(現在の爬虫類の大半はここに含まれます)にまたがっていることも爬虫類を見直す要因の一つになっています。
恐竜は一時代前の生物であり化石が比較的豊富に産出されるので、その生態についてかなり理解が進んでいます。次のような課題については一定の考え方が出ています。
(1) 恐竜は恒温動物か変温動物か
(2) 鳥類の祖先は爬虫類か恐竜か
(3) 羽毛をもった恐竜はいたのか
(4) 恐竜は気嚢システムをもっていたか
(5) 恐竜は2足歩行か4足歩行か
(6) 竜脚類はどのようにして歩行していたか
(7) 最大級の竜脚類はどのくらいの植物を食べていたのか
(8 )ティラノサウルスは俊敏なハンターだったのか
恐竜の中で最大級のものは竜脚類の仲間で体重が100トンにもなります。この体重を維持するためにはどのくらい大量の食物が必要なのか興味のあるところです。というのは竜脚類の仲間は非常に長い首をもっており、その先端に付いている頭部は現在の馬くらいでした。歯も発達しておらず、噛み切るというより噛みちぎって胃の中に送っていたようです。
食料が十分にある環境でも、この構造では(休まずに食べても)1日に1トンくらい食べるのが限界であったと考えられます。現在のアフリカゾウ(体重は5-7トン)は毎日150kgの植物を必要としています。
哺乳類の場合の標準代謝量(安静時の代謝量、基礎代謝)は体重の3/4乗に比例します。もし、体重60トンの竜脚類が哺乳類と同じ基礎代謝をもっていたら、アフリカゾウの6倍くらいの植物を食べなければならない計算となります。これはどうみても非現実的な値ですので、大型恐竜の基礎代謝は哺乳類より相当低かったと推定できます。
さいわい体が大きいと体温は恒温的なります。コップのお湯はすぐ冷めてもお風呂のお湯はなかなか冷めないのと同じ原理です。このような性質を慣性恒温性といいます。小型の恐竜は恒温動物であった可能性は高いのですが、大型恐竜は進化の過程で基礎代謝を下げる省エネルギー生活を身に付けたのでしょう。
それでも、アフリカゾウの2倍程度の食料は必要であったと考えられますので、起きている時間はいつも食べていたのではと推測できます。大きなことはいつも良いこととは限りません。
竜脚類の大きさに関してはもう一つの話題が提供されています。竜脚類は長い首と長い尻尾のバランスをとって歩行していたと考えられますが、あの長い首はどのくらいの高さまで持ち上げられられたのかが議論になっています。映画のジュラシックパークやNHKの恐竜番組では90度近くまで持ち上げる姿になっています。
しかし、動物生理学的な観点からあの首は左右に振って食料を確保するためのもので、上方には20度くらいまでしか持ち上げられなかったという学説が出てきています。真上に持ち上げた状態で頭部に血液を送るために必要な大きくて力強い心臓は持ち得ないという考え方です。
確かに動物生理の観点からは一理ありますが、それでは竜脚類の食料は地上10mくらいのところに限定されてしまい、当時の裸子植物(針葉樹)の葉の大半を食べることができません。もっとも豊富にある植物を食べられないとすると、1日300kgの食べ物は手に入ったのでしょうか。
最強の肉食獣とされるティラノサウルスが俊敏なハンターであったかどうかも議論になっています。あごの強さと肉を噛み切る鋭い歯については文句のないティラノサウルスですが、成体は6-7トンもあり、2足歩行でたとえばトリケラトプスより早く動くことができたのでしょうか。
ティラノサウルスについては死肉をあさるスカベンジャーであったという説も根強くあります。しかし、現在の世界ではスカベンジャーとしての大型肉食獣はいません。獲物がそんなに都合よく死んでくれないからです。さらに、他の死肉をあさる肉食恐竜との競合もあるので巨大なティラノサウルスはスカベンジャーとしてはとてもやっていけなかった考えるのが妥当です。
NHKの恐竜番組ではティラノサウルスは17歳くらいで成熟し、その直前に急速に大きくなるという説が紹介されています。17歳より若い個体は体重が少ないので俊敏であり、彼らが獲物を追いかけ、成体の隠れている場所に追い込むという家族ぐるみの狩りが紹介されていました。確かにこれはありあそうな話です。
ちなみに、アフリカゾウの走る速度は時速40kmですので、同じくらいの体重、体の造りのトリケラトプスもこの程度の速度を出せたと考えられています。ところが、ティラノサウルスは現存するモデルがありませんので、せいぜい時速5kmであったいう説から40kmくらいだという説まで幅があります。
良質な恐竜の化石は日本でも見つかるようになりました。化石が見つかるためには恐竜の時代の地層が地表付近に出ていなければなりません。ようやく、福井県勝山市北谷において恐竜時代の地層の露頭が見つかり、獣脚類やイグアノドン類をはじめとして多くの恐竜化石が発掘されています。
発掘現場からは恐竜の足跡化石も見つかり、たくさんの恐竜の群れが生息していたことが明らかになりました。その縁から福井県には県立恐竜博物館があり、そこのHPには恐竜の復元標本のデータベースがあります。詳細は「 福井県立恐竜博物館 」を訪問してください。
1.5億年にわたり地上を支配してきた恐竜も突然最後のときがやってきます。その原因として広く支持されているのは隕石落下説です。白亜紀末の6500万年前に直径10kmほどの隕石が落下しました。
この隕石が形成したと考えられる直径180kmの巨大なクレーターがユカタン半島で発見されています。また、世界中の白亜紀と古第三紀の境界層には隕石起源の大量のイリジウムが発見されています。
直径10kmの隕石が秒速20kmで当時は浅海だった地表に衝突しました。衝突のエネルギーは広島型原爆の約50億倍に相当し、大気中に拡散した大量のちりと森林火災による煤が太陽光を遮断しました。気温の急激な低下とそれに続く温暖化、光合成を行う植物やプランクトンの死滅などの結果、恐竜をはじめ多くの生物種が絶滅しました。
隕石衝突とそれに続く環境の激変は生物にとっては過酷なものであったことは想像に難くありませんが、両生類、爬虫類、哺乳類、鳥類の一部は生き残り、恐竜だけが絶滅した自然の選択性にはまだまだ謎が残されています。大きさが選択の一つの基準であったことは確かでしょうが、小型の恐竜も絶滅しており、やはり謎は残されます。
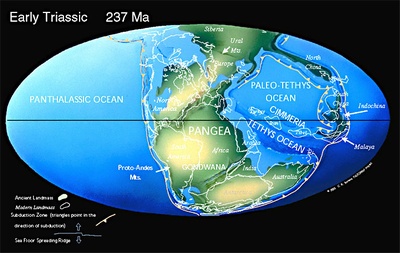
三畳紀の大陸配置、画像は
「PALEOMAP Project」より引用しました。超大陸パンゲアはC字型をしており、赤道付近にはチチス海が広がっています。この時期からパンゲアは分裂し、2.5億年で現在の大陸の配置になっています。
シベリア洪水玄武岩(シベリア・トラップ)、画像はwikipedia より引用しました。この広大な地域にマグマがあふれました。
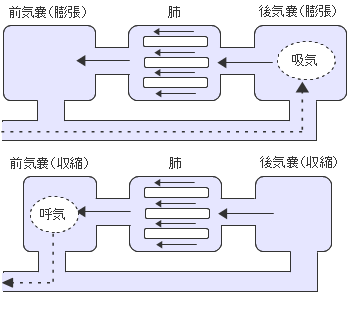
鳥類の気嚢システム、気嚢を膨張・収縮させることにより体内で空気の流れを作ることができます。吸気の時は前気嚢、後気嚢の両方を膨らませ後気嚢に新鮮な空気を取り込みます。呼気の時は両方の気嚢を収縮させて肺から出た空気を外に出します。この仕組みにより肺にはいつも新鮮な空気が送り込まれ、ガス交換の済んだ空気が排出される一方向の流れができています。
| 区分 | 種類 | 体長(m) | 体重 |
|---|---|---|---|
| 恐竜 | プラキオサウルス(竜脚類) アパトサウルス(竜脚類) アロサウルス(獣脚類) ティラノサウルス(獣脚類) ステゴサウルス(剣竜類) トリケラトプス(角竜類) ミクロラプトル(獣脚類) ヴェロキラプトル(獣脚類) イクチオサウルス(魚竜) プレシオサウルス(首長竜) プテラノドン(翼竜) |
24-28 22-23 8-10 13 7-9 8-10 0.4 2 2 2-5 翼長9m |
30-80 ton 25-30 ton 3-4 ton 6-7 ton 3-7 ton 3-8 ton 2-3 kg 15 kg 250 kg 0.5-1 ton 25 kg |
| 恐竜以外 | シロナガスクジラ アフリカゾウ ライオン タイリクオオカミ ガラパゴスゾウガメ ダチョウ ジンベイザメ クロマグロ |
20-30 6-7.5 1.5-2.5 1.5 1.5 2 10-13 3 |
150 ton 5-7.5 ton 150-250 kg 50 kg 250 kg 90 kg 18 ton 400 kg |
生物の大きさ比較、この表から分かるように恐竜の仲間は他の脊椎動物に比べて非常に大型であったことが分かります。もっとも現在の哺乳類がそうであるように、恐竜の中にもニワトリくらいの大きさのものもいました。
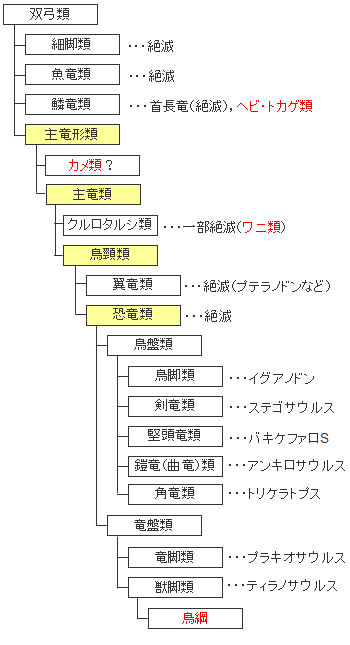
化石動物を含めた双弓類の簡易系統図、学術的に恐竜にに分類されるのは陸上のものだけです。翼手竜、魚竜、首長竜などは恐竜には含まれません。鳥類は恐竜類の唯一の生き残りの種ということになります。ワニ類は恐竜にもっとも近い爬虫類ということになります。

ティラノサウルスとディロング、画像は「古代世界の住人・ブログ」から引用しました。このサイトには名前の通りあらゆる古代生物の画像が用意されています。ブログにはディロングとティラノサウルスが一緒に出ていたので引用しました。ディロングは白亜紀の前期に現れた体長1.5mほどの羽毛恐竜であり、ティラノサウルスの祖先と考えられています。ティラノサウルスも若い個体は羽毛をもっていたと考えられます。成体は慣性恒温性になりますので、逆に体温の上昇を抑えるため羽毛を失ったようです。
哺乳類の時代(6500万年前~)
1.5億年にもわたる恐竜の時代にも哺乳類の祖先は存在していました。大きさはネズミほどで夜行性であったと考えられています。昼間活動し豊富な食料に恵まれていた恐竜は巨大化への道を歩みましたが、その間、哺乳類は少なくとも外観上はさしたる進化をしていません。
しかし、恐竜の時代の終わり頃には小さな犬ほどの哺乳類が恐竜の子どもを食べていたことが分かっています。また、夜行性の哺乳類は聴覚、さらには脳を発達させ敏捷な運動機能も手に入れました。
約6500万年前に恐竜が絶滅するとその空白を埋めるように哺乳類は爆発的に進化し、多種多様な種が現れました。恐竜という最大の競争種が絶滅したことにより、哺乳類が新しい時代の最強種となり適応放散していった結果であると考えられています。
草食のもの、肉食のもの、大きなもの、小さいなもの、樹上で暮らすもの、水中で暮らすもの、空を飛ぶもの…、哺乳類の世界はその前の恐竜の世界と非常に類似しています。そして、それぞれの環境に適応した生物は哺乳類、爬虫類、恐竜という種の違いはあっても、その体のデザインは驚くほど似通っています。
このように種は異なっているにもかかわらず、同じような環境に暮らしている生物の体型や特定の器官がとてもよく類似することを収斂進化といいます。生物は環境に適応したものが生き残る可能性が高いというのは自然界の厳粛な掟であるため、異なる祖先から出発しても同じような環境で同じように生きている生物同士は互いに似てくるということです。
カンブリア爆発の研究者であるサイモン・モリスはその著書「カンブリア紀の怪物たち」の中で「ある生物の系統における予想される進化の最終産物の形態はだいぶ限定されるように思われる。進化は拘束されない無制限の実験の連続であるということはできない。むしろ、一定の範囲内では進化のプロセスの結果は予測可能であると考えるべきである」と記しています。
大絶滅は何回か発生しました
多くの場合、環境の変化は生物進化のトリガーになります。しかし、生物の適応能力を超える大変化があった場合、多くの生物種は生き延びることができませんでした。生物種の過半数あるいは95%が死滅するという大絶滅は地球史において何回か発生しています。
そのような大絶滅は環境の激変により引き起こされたものですが、どのようなイベントがどのような環境の変化をもたらしたかについては定説はありません。もっとも近い6500万年前の「K-T境界の大絶滅」については隕石衝突説が定説になっていますが、一つの事実の発見がこの定説をくつがえす可能性は大いにあります。
なんといっても6500万年も前の事件ですから原因を正確に把握することは困難です。多くの事実は化石よりもたらされます。しかし、発見される化石は非常に少ないのも事実です。1.5億年の歴史をもっている恐竜にしても、その50%の種はただ1点の化石標本から決められています。恐竜が化石になり私たちに発見される確率はそのくらい低いことになります。そのような化石で6500万年前の真実の証拠を見つけ出すのは至難の業ということができます。
これがさらに古い時代となると大絶滅の原因を調べることはますます困難となります。2.5億年前のペルム紀末の大絶滅(P-T境界)の原因は「シベリア・トラップの形成」、つまり大規模な火山活動によるものという仮説が有力ですが、まだ定説にはなっていません。
現在、地球上にはどのくらいの生物種が存在するかは正確には分かっていません。およそ400万種から4000万種と見られていますが、人類が分類した生物種はこれまで180万種に過ぎません。人類だけが非常な勢いで人口を増加させ、地球上のあらゆる場所に進出している一方で、生息場所を狭められたり、乱獲により絶滅の危機に瀕している生物種も少なくありません。
人類がこのまま他の生物を圧迫し続けるならば、現代は「人類が引き起こした大絶滅の時代」と呼ばれることになるでしょう。40億年にわたる生命の歴史を今に伝える生物の多様性を次の時代に引き継いでいくのは人類の責務です。
| 発生時期 | 名称 |
|---|---|
| 5.7億年前 | 原生代末期の大絶滅(V-C境界) |
| 4.4億年前 | オルドビス紀の大絶滅 |
| 3.7億年前 | デボン紀後期の大絶滅 |
| 2.5億年前 | ベルム紀末の大絶滅(P-T境界) |
| 6500万年前 | 白亜紀末の大絶滅(K-T境界) |
生物種の過半数あるいは95%が死滅するという大絶滅は地球史において何回か発生しています。そのような大絶滅は環境の激変により引き起こされたものですが、どのようなイベントがどのような環境の変化をもたらしたかについては定説はありません。
参照サイト
- 海と船なるほど豆事典
- 共通祖先(wikipedia)
- 原始生命体核生物(wikipedia)
- 生命の起源(wikipedia)
- 独立栄養生物(wikipedia)
- 原核生物(wikipedia)
- 真核生物(wikipedia)
- 細胞内共生説(wikipedia)
- 共生(wikipedia)
- 縞状鉄鉱床(wikipedia)
- エディアカラ生物群(wikipedia)
- カンブリア紀(wikipedia)
- オゾン層(wikipedia)
- 石炭紀(wikipedia)
- 爬虫類(wikipedia)
- 生物の起源
- NHKかがく用語集
- 生物の進化
- 真核生物誕生の謎
- 古代世界の住人
- 啓林館・動物の分類と系統
- 動物大きさ比べ
- 恐竜の謎
- 福井県立恐竜博物館
- 空気の基礎知識